Учебное пособие Часть I самара Самарский государственный технический университет 2007
 Скачать 1.81 Mb. Скачать 1.81 Mb.
|

|
Образование гидантоинов. При нагревании α-аминокислот с мочевиной без растворителя образуются гидантоины. При обработке водных или неводных растворов α-аминокислот изоцианатами в зависимости от условий могут быть получены гидантоиновые кислоты или гидантоины:  Гидантоины    Гидантоины Гидантоиновые кислоты Гидантоины часто используют для идентификации α-аминокис-лот, поскольку они легко кристаллизуются и имеют четкие температуры плавления в отличие от аминокислот. Образование дикетопиперазинов. Как было описано ранее, α-аминокислоты, а ещё легче их эфиры, циклизуются с образованием 2,5-дикетопиперазинов. Например, простым нагреванием глицина в этиленгликоле при 174-176ºС с выходом 62% был получен 2,5-дикетопиперазин, а метиловый и этиловый эфиры глицина спонтанно переходят в 2,5-дикетопиперазин в водном растворе при 15-20ºС. Рассмотренные ранее реакции образования из α-аминокислот азлактонов, тиогидантоинов и реакции α-аминокислот с нингидрином и N-бромсукцинимидом также идут с участием и амино– и карбоксильной групп. Однако авторам представлялось более логичным рассмотреть их в п. 1.1.4.2, поскольку начинаются они с участия аминогруппы, а заканчиваться могут в зависимости от условий и без участия карбоксильной группы. Правка отсюда! 1.1.4.4. Специфические реакции аминокислот Аминокислоты, содержащие реакционноспособные боковые цепи, могут принимать участие в реакциях, специфических для этих цепей. Специфические реакции представляют интерес по двум причинам. Во-первых, реакции, сопровождающиеся характерными проявлениями, широко используются для идентификации и количественного определения пептидов, белков и индивидуальных аминокислот. Во-вторых, реакции, характерные для боковых цепей аминокислот, часто используются для химической модификации пептидов и белков. Рассмотрим специфические реакции аминокислот, имеющие аналитическое значение (табл.1.5). Ксантопротеиновая реакция. Аминокислоты с ароматическими кольцами в боковой цепи, а также пептиды и белки, в состав которых входят такие аминокислоты, при нагревании с концентрированной азотной кислотой дают желтое окрашивание, которое обусловлено образованием нитросоединений. Нитросоединения тирозина и триптофана в отличие от нитрофенилаланина при подщелачивании образуют аци-нитросоли оранжево-красного цвета, что позволяет отличить фенилаланин от тирозина и триптофана:  Таблица 1.5 Специфические реакции, используемые для идентификации и количественного анализа α-аминокислот и белков
Как видно из сущности ксантопротеиновой реакции, определению тирозина, триптофана и фенилаланина мешают ароматические соединения, и прежде всего фенолы и ароматические амины. Реакция Миллона. Этот метод определения тирозина основан на том, что, во-первых, тирозин в отличие от триптофана и фенилаланина нитруется даже разбавленной азотной кислотой, а во-вторых, образующийся нитротирозин дает красную, нерастворимую в кислой среде аци-нитросоль с ионом [Hg2]2+:  При низкой концентрации тирозина возникает красное окрашивание, при высокой – выпадает красный осадок. Эта реакция положительна также для фенольных соединений. Нитропруссид натрия – Na2[Fe(CN)5NO] дает с меркаптанами в щелочной среде характерную красную или красно-фиолетовую окраску вследствие образования комплексных соединений следующего типа: N Цистеин дает положительную реакцию в разбавленном растворе аммиака, а цистин и метионин – отрицательную. Аммиак, а не щелочь, как обычно, используется потому, чтобы исключить возможность образования меркаптанов из цистина и метионина, меркаптаны дают положительную реакцию с нитропруссидом натрия. Реакция Фоля. Метод Фоля позволяет определять наличие серы в меркаптанах и сульфидах. Метод заключается в кипячении раствора препарата с щелочным раствором плюмбита натрия; при этом образуется черный осадок сульфида свинца. Положительную реакцию Фоля дают не только цистеин, цистин и метионин, но и пептиды и белки, содержащие эти аминокислоты: Определению серосодержащих аминокислот мешают соединения, содержащие в своей молекуле сульфидную или меркаптановую функциональные группы, например антибиотик пенициллин. Реакция Сакагучи. Аргинин легко конденсируется с α-дикетонами с образованием имидазольного цикла, что используется в пептидном синтезе для защиты гуанидиновой группы аргинина. В реакции Сакагучи происходит аналогичная конденсация с 1,2-нафтохиноном, который образуется при окислении α-нафтола гипобромитом или гипохлоритом натрия. Реакция Сакагучи является общей для любых производных гуанидина. Так, например, антибиотик стрептомицин также дает положительную реакцию Сакагучи, поскольку содержит в своей молекуле гуанидиновый остаток ((H2N)HN=C-NH-):  Открытие глутаминовой кислоты и глутамина. При нагревании глутаминовой кислоты или глутамина они циклизуются с отщеплением воды и образованием пирролидонкарбоновой кислоты (пироглутаминовая кислота) или ее амида соответственно:  Пироглутаминовая кислота при нагревании с резорцином в присутствии концентрированной серной кислоты образует краситель, приобретающий в щелочной среде красно-фиолетовый цвет:   Реакция Паули. Эта реакция основана на азосочетании активированных ароматических соединений с солями диазония, в результате которой образуются азокрасители. Соль диазония готовят из сульфаниловой кислоты:   Реакция Фолина-Чиокалтеу. Среди протеиногенных аминокислот тирозин является самым сильным восстановителем. На использовании этого свойства и основана реакция Фолина-Чиокалтеу. Из всех протеиногенных аминокислот реактив Фолина-Чиокалтеу, представляющий собой фосфомолибдовольфрамовую кислоту, окисляет лишь тирозин по схеме  При этом фосфомолибдовольфрамовая кислота восстанавливается до фосфомолибдовольфрамовой гетерополисини – вещества непостоянного состава синего цвета. 1.2. Полипептиды 1.2.1. Природа пептидной связи «Я предложил название «полипептид» для продуктов, образуемых при соединении аминокислот связями амидного типа. Простейшим представителем этого класса соединений является гликоколь, так называемый глицилглицин, H2NCH2CO-NHCH2COOH. По количеству аминокислот, входящих в состав пептида, различают ди-, три-, тетрапептиды и т.д.». Так Эмиль Фишер определил пептидную связь как амидную связь между аминокислотами (E. Fischer, Ber., Bd. 39, S. 530-610, 1906 г.). Основная структурная особенность полипептидов – наличие цепей, составленных из аминокислотных остатков, связанных между собой α-амидными связями:  В биохимии эти связи принято называть пептидными. Фундаментальная концепция, принятая еще в 60-х гг. прошлого века во всех работах, посвященных изучению конформации полипептидов, заключается в планарности транс– и цис-пептидной групп. Этот факт указывает на наличие n-π сопряжения между неподеленной парой электронов атома азота (n) и π-электронами карбонильной группы, что подтверждается также меньшей длиной связи между атомом азота и атомом углерода карбонильной группы (C−N = 1,32 Å) по сравнению с длиной связи между этим же атомом азота и α-углеродным атомом (N−Cα = 1,47 Å): 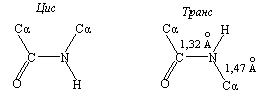 Вследствие n-π сопряжения вращение вокруг связи C–N затруднено. Поворот вокруг этой связи обозначается торсионным углом ω. Обычно угол ω равен 180° (транс-пептидная связь), однако изучение конформаций полипептидов показало что в некоторых случаях наблюдается искажение планарной формы отдельных пептидных звеньев (так называемая «пирамидальность») в пределах ±15°. Например, в кристаллической структуре гидрохлорида глицил-L-аланина отклонение от плоскости составляет +10,2º, а для глицил-L-лейцина -11,4º. В отличие от этого вращение вокруг связей Сα−С и N−Cα осуществляется свободно и характеризуется двугранными торсионными углами ψ и φ соответственно, однако это не значит, что торсионные углы ψ и φ могут принимать какие угодно значения. Для каждого конкретного сочетания определенных аминокислотных остатков ввиду стерических ограничений разрешены определенные комбинации торсионных углов φ и ψ. Для углов φ и ψ остаются разрешенными сочетания, лежащие в пределах определенных дискретных областей. Информацию о связи между торсионными углами φ и ψ в каждом пептидном звене представляют графически с помощью конформационной φ/ψ-карты (карты Рамачандрана). На рис.1.6 представлена карта, отражающая возможные энергетически выгодные, энергетически невыгодные, но возможные, и запрещенные сочетания торсионных углов φ и ψ. Как видно из рис. 1.6, в полипептидах реализуются весьма ограниченные соотношения торсионных углов φ и ψ. Определенные возможные сочетания углов, как правило, соответствуют определенным упорядоченным конформациям участков полипептидной цепи (подробнее о структурной организации полипептидов см. п. 1.2.2.3.1). На рис. 1.6 цифрами отмечены области углов φ и ψ, характерные для разных типов вторичной структуры белков. 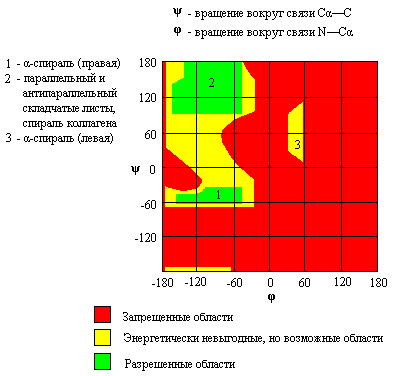 Р и с.1.6. Конформационная φ/ψ – карта полипептидной цепи 1.2.2. Классификация полипептидов, отдельные представили и их биологическая роль С развитием биохимии термин «полипептиды», введенный Э.Фишером, приобрел более широкое значение. В настоящее время под полипептидами понимают соединения, состоящие из аминокислот, связанных между собой не только пептидными связями, но и связями других типов, а также содержащие в своей структуре не только протеиногенные аминокислоты, но и не встречающиеся в белках. Все полипептиды по химическому строению делятся на следующие группы:  1.2.2.1. Малые линейные пептиды К этой группе относятся полипептиды, содержащие до 50 аминокислотных остатков и имеющие молекулярную массу не более 5000 Да. Малые линейные пептиды делятся на низкомолекулярные (до 10-12 аминокислотных остатков в цепи) и высокомолекулярные (12-50 аминокислотных остатков в цепи). Крупные полипептиды на основе α-аминокислот, связанных между собой пептидными связями, с молекулярной массой более 5000 Да называют белками. Естественно, что четкой границы между высокомолекулярными пептидами и белками нет. Малые линейные пептиды подразделяют на гомомерные и гетеромерные. Гомомерные линейные пептиды построены исключительно из α-L-аминокислот и содержат только α-пептидную связь (белковоподобные пептиды). Малые гомомерные линейные пептиды в природе встречаются так же часто, как и белки, и прежде всего в организмах животных. В качестве примера рассмотрим важнейшие представители этой группы. По биологическим свойствам эта группа пептидов делится на три основные группы: нейромедиаторы, пептидные гормоны, нейротоксины некоторых животных. Нейромедиаторы. В 1975 г. из мозга млекопитающих были выделены два природных опиоидных пентапептида – [Met5]-энкефалин и .[Leu5]-энкефалин, воздействующие на опиатные рецепторы, но в отличие от морфина, не обладающие эффектом привыкания: Tyr– Gly-Gly-Phe-Met – [Met5]-энкефалин Tyr– Gly-Gly-Phe-Leu – .[Leu5]-энкефалин Впоследствии из тканей гипофиза и гипоталамуса млекопитающих были выделены и другие полипептиды с аналогичной биологической активностью. Такие полипептиды, как природные, так и синтетические, получили название опиоидные полипептиды. Все они характеризуются присутствием остатка [Met5]-энкефалина в конце цепи (выделен жирным шрифтом). Их действие обусловлено способностью связываться с опиатными рецепторами вследствие сходности пространственного строения энкефалинового фрагмента и морфина. Общее название эндогенных полипептидов, обладающих опиоидной активностью, – эндорфины. Ниже приведены формулы выделенных и охарактеризованных эндорфинов человека: α-эндорфин – Tyr-Gly-Gly-Phe-Met-Thr-Ser-Glu-Lys-Ser-Gln-Thr-Pro-Leu– Val-Thr γ-эндорфин – Tyr-Gly-Gly-Phe-Met-Thr-Ser-Glu-Lys-Ser-Gln-Thr-Pro-Leu- Val-Thr-Leu δ-эндорфин – Tyr-Gly-Gly-Phe-Met-Thr-Ser-Glu-Lys-Ser-Gln-Thr-Pro-Leu- Val-Thr-Leu-Phe-Lys-Asn-Ala-Ile-Ile-Lys-Asn-Ala-Tyr β-эндорфин – Tyr-Gly-Gly-Phe-Met-Thr-Ser-Glu-Lys-Ser-Gln-Thr-Pro-Leu- Val-Thr-Leu-Phe-Lys-Asn-Ala-Ile-Ile-Lys-Asn-Ala-Tyr-Lys-Lys- Gly-Gln – Энкефалины и эндорфины блокируют передачу нервных импульсов, поступающих от внутренних органов в подсознание, из подсознания – в сознание. Именно поэтому здоровый человек не ощущает пульсацию внутренних органов в том случае, если уровень поступающих от них импульсов не превышает определенную норму. Морфин и другие наркотики, воздействуя на опиатные рецепторы, выполняют ту же функцию, что и природные эндорфины и энкефалины, однако при этом развивается привыкание, биохимическая основа которого заключается в том, что организм перестает вырабатывать природные эндорфины, и поэтому нуждается в постоянном приеме наркотика. Попытка прекратить прием наркотика вызывает состояние «ломки», когда импульсы, поступающие в подсознание от всех внутренних органов, беспрепятственно поступают и в сознание, что ощущается как чувство сильнейшей боли, ведь выработка природных эндорфинов и энкефалинов организмом прекращена. Все было бы не так страшно, если бы наркотики не дезорганизовывали работу внутренних органов. Вот почему наркоманы долго не живут. Чтобы излечиться от наркомании, необходимо восстановить продукцию естественных энкефалинов и эндорфинов, которые, по-видимому, образуются в центральной нервной системе из специфического белка под действием специфических пептидаз. Процесс этот длительный и далеко не комфортный, вот почему так трудно избавиться от наркотической зависимости. Пептидные гормоны. Гормоны животных (от греч. hormao – приводить в движение, побуждать), вещества, вырабатываемые специализированными клетками и железами внутренней секреции и регулирующие обмен веществ в отдельных органах и всем организме в целом. Органы, в которых вырабатываются пептидные гормоны животных, – это поджелудочная железа, желудок, гипофиз, почки, кровь. Ниже приведены формулы некоторых не очень больших гомомерных линейных пептидных гормонов, а в табл. 1.6 – краткие сведения об их биологическом действии. Ангиотензин I – Asp-Аrg-Val-Tyr-Ile-His-Pro-Phe-His-Leu Ангиотензин II – – Asp-Аrg-Val-Tyr-Ile-His-Pro-Phe Брадикинин – Arg-Pro-Pro-Gly-Phe-Ser-Pro-Phe-Arg Глюкагон – His-Ser_Gln-Gly-Thr-Phe-Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-Leu-Asp- Ser-Arg-Arg-Ala-Gln-Asp-Phe-Val-Gln-Try-Leu-Met-Asn-Thr Меланофорстимулирующий гормон – β-МСГ – Ala-Glu-Lys-Lys-Asp-Glu-Gly-Pro-Tyr-Arg-Met-Glu-His- Phe-Arg-Tre-Gly-Ser-Pro-Pro-Lys-Asp В табл. 1.6 представлены не только гомомерные линейные пептидные гормоны, но и представители других типов пептидных гормонов – гетеромерные и гетеродетные пептидные гормоны. Нейротоксины животных. Нервные яды пресмыкающихся, ядовитых насекомых, морских червей и других животных представляют собой главным образом белки. Однако известны и гомомерные пептидные яды. Например, яды пчел и ос представляют собой сложные смеси полипептидов, основным компонентом которых является гомомерный пептид – мелиттин (его содержание достигает 50%): Gly-Ile-Gly-Ala-Val-Leu-Lys-Val-Leu-Thr-Thr-Gly Leu-Pro-Ala-Leu- -Ile Ser-Trp-Ile-Lys-Arg-Lys-Arg-Lys-Arg-Glu-Glu Молекула мелиттина состоит из 28 аминокислотных остатков. Примечательно, что молекула мелиттина является гомомерной и не содержит цистеина, в отличие от нейротоксинов змей, скорпионов, ядовитых пауков, которые являются гетеродетными пептидами и, как правило, содержат несколько дисульфидных мостиков. Таблица 1.6 Пептидные гормоны
|