Ответы на билеты по биологии. Билет1 Закономерности существования клетки во времени. Жизненный цикл клетки, его варианты. Основное содержание и значение периодов жизненного цикла клетки
 Скачать 2.17 Mb. Скачать 2.17 Mb.
|

|
Первичное – соотношение полов на уровне ранних эмбрионов (100 женских на 140 мужских). Вторичное – соотношение полов на момент рождения (100 девочек:107 мальчиков). К моменту вступления в брак (18-20 лет) на 100 женщин приходится 101 мужчина. К 50-ти годам – на 100 женщин – 85 мужчин. Половые различия особей обусловлены первичными и вторичными половыми признаками. Кроме того, пол характеризуется признаками: зависящими от пола, ограниченными полом и сцепленными с полом. Первичные половые признаки – признаки, которые обеспечивают непосредственно процесс образования половых клеток, оплодотворения и развития потомства. К ним относятся внутренние и наружные половые органы. Формирование обусловлено генами, расположенными в половых хромосомах. Зачатки гонад закладываются на поверхности первичных почек из материала мезодермы. Сюда мигрируют первичные половые клетки, появляющиеся на стадии гаструляции в энтодерме желточного мешка. Начинается анатомическое формирование – гонадогенез. Гонады первоначально закладываются одинаково у обоих полов. В ходе дифференцировки пола идет подавление одного слоя гонады и развитие другого (кортекса над медуллой). У мужского пола быстрее развивается медуллярная ткань, а у женского – кортекс. Семенники формируются на 6-12 неделе, яичники на 14-16 неделе внутриутробного развития. Вторичные половые признаки – признаки, которые не принимают непосредственного участия в воспроизводстве потомства, но связанны с ним и косвенным образом влияют на него. Сюда относятся фенотипические различия между мужчинами и женщинами. Например, наличие/отсутствие молочных желез, рост, масса тела, тип телосложения, тип оволосения, тембр голоса. Также обусловлены генами половых хромосом и генами аутосом. Признаки, ограниченные полом – признаки, гены которых имеются в кариотипе обоих полов, но проявляются преимущественно лишь у одного пола. Такие анатомические и физиологические черты, как развитие молочных желез, время начала менструации, особый тип телосложения контролируются генами, получаемыми от обоих родителей, но проявляются лишь у женского пола. Сугубо мужские черты, как характер роста волос на лице, распределение волосяного покрова на лице также контролируются генами, общими для обоих полов. Признаки, зависящие от пола – признаки, определяющиеся генами, которые есть у обоих полов, но у одного они ведут себя как доминантные, а у другого – как рецессивные. Например, ген облысения – ген аутосомный. Признаки, сцепленные с полом – неполовые признаки, гены которых локализованы в половых хромосомах. У человека Х хромосома крупная субметацентрическая и относится к группе С по денверской классификации. Х-хромосома человека богата жизненно важными генами, которые контролируют свертываемость крови, способность различать цвета, гены, контролирующие мышечный тонус, проницаемость стенки сосудов и другие. У хромосома человека имеет небольшие размеры, это самая мелкая хромосома в кариотипе человека, бедна жизненно важными генами (относится к группе G). К таким генам относятся общая цветовая слепота, Пигментная ксеродерма, Пигментный ретинит, Геморрагический диатез, болезнь Огучи. Гены, полностью сцепленные с Х хромосомой, могут быть как доминантными, так и рецессивными. Х-сцепленный доминантный тип – фолликулярный гиперкератоз, наследственный рахит, потемнение зубной эмали. Рецессивные признаки – дальтонизм, атрофия зрительного нерва, несахарный диабет, ихтиоз, отсутствие потовых желез. Наиболее известный пример – гемофилия. Из наиболее редких и уникальных – синдром Леша-Найхана (самосадизм). Такие больные характеризуются тяжелыми неврологическими расстройствами, сниженным болевым порогом, неудержимым стремлением к самоповреждению. Рецессивные гены, полностью сцепленные с Х хромосомой, наследуются то типу крисс-кросс (от отца через дочь-носительницу к внукам мужского пола). Закономерности наследования:
Согласно гипотезе Лайон, в нормальных женских клетках функционирует одна из двух Х хромосом, а другая подвергается инактивации (на 16 сутки эмбриогенеза). Это является механизмом дозовой компенсации генов. В одних клетках женского организма инактивации подвергается отцовская Х хромосома, в других – материнская. Инактивированная Х хромосома определяется в виде мелкой глыбки (тельце бара, половой хроматин). В нормальных женских клетках 1 тельце Барра, в нормальных мужских – отсутствует. Исследование полового хроматина (может быть с помощью буккального соскоба) проводится для определения пола эмбриона, в диагностике половых генетических аномалий. Половые генетические аномалии являются следствием нерасхождения половых хромосом в мейозе. Это происходит в 0,3% всех гамет. В яйцеклетке вместо одной Х хромосомы может оказаться две или не будет ни одной. При оплодотворении таких аномальных яйцеклеток нормальными сперматозоидами будут образовываться зиготы, в которых изменено количество половых хромосом. Возможны отклонения и при сперматогенезе (УО). Аномалии количества половых хромосом могут быть в виде моносомий и полисомий. Трисомия Х. Кариотип ХХХ. Рождается девочка (1:1000). Фенотипические проявления разнообразны. Большинство женщин имеют ряд нерезких отклонений в физическом развитии, нарушении функции яичников, преждевременный климакс, интеллектуальную неполноценность. Но у некоторых больных могут не проявляться. В клетках наблюдаются два тельца полового хроматина. Синдром Шерешевского-Тернера. Моносомия Х. Кариотип ХО. Фенотип женский. 1:4000-1:5000. Основной патологический признак – недоразвитие яичников, они недифференцированы и представляют собой зачатковые тяжи, в которых почти нет фолликулов. Отсутствие гонад ведет к появлению женского пола по фенотипу, но половые различия сглажены. Характерна диспропорция тела, нижние конечности укорочены, широкие плечи, узкий таз, рост 135-145 см. Короткая шея со складками кожи, идущими от затылка (шея сфинкса), низкий рост волос на затылке, антимонголоидный разрез глаз. Умственное развитие не страдает. Отмечается некоторая инфантильность, неустойчивость настроения. Половой хроматин в клетках отсутствует. Такие женщины бесплодны. Синдром Клайнфельтера. ХХУ (м.б. ХХХУ, ХХХХУ). Мужской фенотип. 1:1000. Эти больные достаточно жизнеспособны. Характерная особенность – недоразвитие семенников, отсутствие сперматогенеза. Развивается астенический тип телосложения, узкие плечи, широкий таз, жироотложение по женскому типу, скудная растительность на лице или полное отсутствие. Наблюдается умственная отсталость, проявляющаяся в разной степени (при избыточном числе Х хромосом – более глубокие нарушения физического и психического развития.) Изменения в фенотипе во время соматического развития, связанные с присутствием в организме генетически разнородных клеток – мозаицизм. Проявляется в виде секторов нового фенотипа, находящихся внутри ткани исходного фенотипа. ХХХ/ХХ – иногда легкая олигофрения, непостоянные нарушения функции гонад. ХО/ХХ и ХО/ХХХ – синдром Тернера, очень разные степени проявления. ХХ/ХУ = истинные гермафродиты. ХХУ/ХУ и ХХУ/ХХ – сходен с синдромом Клайнфельтера, но с более сглаженными симптомами. Гермафродитизм.
При истинном гермафродитизме гонады являются неполноценными анатомически и функционально. Содержат и семенные канальцы, и фолликулы. Возможен вариант, когда с одной стороны яичко, с другой – яичник. Характеризуется женским набором половых хромосом (ХХ), иногда встречаются мозаики (46 ХХ/46 ХУ). Вторичные половые признаки имеют элементы обоих полов. Одним из важных лечебных мероприятий является хирургическое изменение пола. Перемена пола у детей 3—4 лет не вызывает психологических трудностей. В возрасте 4—10 лет такая перемена переносится тяжело, т.к. обычно в этом возрасте твердо устанавливается сознание половой принадлежности, а понятия пола в смысле половой функции еще не существует. Начиная с пубертатного периода дети осознают свою интерсексуальность и становятся еще более ранимыми. Нередко они сами настаивают на определении пола. В юношеском возрасте (16—17 лет) эта проблема осложняется в связи с появлением тщательно скрываемых и подавляемых половых влечений, обычно гетеросексуальных по отношению к тому полу, в принадлежности к которому воспитывался больной. В целом, у больных в возрасте 16—20 лет адаптация к перемене женского пола на мужской психологически происходит легче, чем при перемене мужского на женский, т.к. особенности мужского поведения приобретаются легче. Смена пола в более старшем возрасте затруднительна по социальным причинам (приобретение определенной специальности, общественного положения и т.п.) При выборе пола главным образом учитывают функциональные особенности наружных половых органов и функциональное превалирование женской или мужской части двуполой гонады. Элементы гонады противоположной избранному полу, по возможности удаляют. Производят хирургическую коррекцию наружных половых органов. Гипогонадизм. Недоразвитость внутренних и наружных половых органов и нечеткая выраженность вторичных половых признаков. Причины множественны (генетически обусловлены, после травмы, инфекционно-воспалительного процесса, в процессе эмбриогенеза, следствие опухоли, кастрации). Тестикулярная феминизация. Женский фенотип, мужской кариотип, мужские гонады. Вместо матки – остатки мюллеровых протоков, вместо фаллопиевых труб – мышечно-волокнистый тяж. Влагалище укорочены, заканчивается слепым мешком. Аменорея. Семенники – в больших половых губах, паховом канале, брюшной полости. Сперматогенез обычно отсутствует. Объясняется ненормальной конечной реакцией органов на гормон, дефектом рецепторов андрогенов. Формирование пола – одно из важнейших аспектов его развития. 4 уровня половой дифференцировки:
3). Экология круглых червей. Морфология, жизненный цикл, патогенность власоглава. Диагностика и профилактика трихоцефалёза. Тип:Круглые черви (Nemathelminthes) Класс:Собственно круглые черви (Nematoda) Вид Заболевание Ascaris lumbricoides (аскарида человеч.) аскаридоз Toxocara canis (аскарида собачья) токсокароз Enterobius vermicularis (острица) энтеробиоз Trichocephalus trichiurus (власоглав) трихоцефалёз Trichinella spiralis (трихинелла) трихинеллёз Strongyloides stercoralis(угрица кишеч.) стронгилоидоз Ancylostoma duodenale(кривоголовка) анкилостомидоз Necator americanus (некатор) некатороз Dracunculus medinensis(ришта) дракункулёз Dirofilaria repens (дирофилярия) дирофиляриоз Filaria(филярии) филяриозы Wuchereria bancrofti (вухерерия) вухерериоз Onchocerca volvulus (онхоцерка) онхоцеркоз Brugia malayi(бругия) бругиоз Согласно классификации В.А.Догеля, тип Nemathelminthes включает в себя 5 классов: 1.Класс Nematoda (Нематоды) 2.Класс Gastrotricha (Брюхоресничные) 3.Класс Gordicea (Волосатики) 4.Класс Kinorinchi (Киноринхи) 5.Класс Rotatoria (Коловратки) Трехслойные нечленистые животные, имеющие первичную полость тела, заполненную жидкостью. Активно передвигаются. Обитают в воде, на почве, во мху. Свободноживущие организмы (коловратки) питаются бактериями, простейшими, водорослями, сами же служат пищей для мальков и взрослых рыб, рачков. Паразитические формы (аскариды, острицы, власоглавы, скребни) живут в организме человека, млекопитающих животных, птиц, рыб, насекомых, ракообразных, которые являются основными или промежуточными хозяевами, вызывая тяжелые болезни, даже массовую гибель. Нематоды паразитируют также в теле растений и грибов, вызывая болезни и снижая урожай. Сельскохозяйственные растения поражаются почвенными нематодами, которые, внедряясь в корни, вызывают их отмирание, образование галлов (вздутий) на стеблях, листьях, клубнях, растрескивание донца у луковичных культур. Поражению нематодами подвержены злаки, овощные культуры, картофель, земляника. Trichocephalus trichiurus (власоглав) - возбудитель трихоцефалёза. Морфология: Передний конец тела червя вытянут в длинный волосовидный придаток. Длина достигает 4-5см, у самок задний расширенный отдел тела изогнут в виде дуги. В нём располагается кишечник и матка. У самцов он закручен в спираль. Власоглавы локализуются в слепой кишке, червеобразном отростке, начальных отделах толстой кишки. Яйца власоглава имеют желтовато-коричневый цвет, по форме напоминают лимон или бочонок. Жизнеспособность яиц 1.5-2 года. Жизненный цикл: Яйца выделяются с фекалиями больного человека. Во влажной почве, при температуре 15-37градусов, яйцо становится жизнеспособным через 1-4 месяца. Рассеянные в почве яйца попадают на овощи, фрукты. Заражение происходит алиментарным путём через грязные продукты и руки. Власоглав развивается без промежуточного хозяина, паразитирует в толстом кишечнике. Вылупившиеся личинки обладают стилетом, они проникают в ворсинки слизистой, где находятся 10 суток. Затем личинки выходят в просвет кишечника, достигают слепой кишки и прикрепляются к слизистой. Половой зрелости они достигают через месяц после заражения. В кишечнике половозрелые формы обитают около 5 лет. Патогенность: Трихоцефалёз может протекать бессимптомно. Серьёзные нарушения работы пищеварительного тракта, сопровождающиеся тошнотой, болями, понижением кислотности. Власоглавы питаются кровью, вызывая развитие анемии. Наблюдается реакция со стороны нервной системы: головная боль, головокружение, припадки. Диагностика: основывается на копрологических исследованиях фекалий. Профилактика: Личная 1.Мытьё рук после посещения улицы, перед едой, после посещения туалета и работы на садовых и приусадебных участках. 2.Тщательное обмывание овощей и фруктов сначала холодной, затем кипячёной водой. 3.Борьба с переносчиками яиц - мухами и тараканами Общественная 1.Санитарно-просветительская работа, направленная на разъяснение населению путей заражения и мер профилактики трихоцефалёза. 2. Своевременное выявление и лечение больных трихоцефалёзом. 3.Предотвращение фекального заражения почвы, благоустройство туалетов. 4.Обезвреживание нечистот, идущих на удобрение огородов, и дегельминтизация почвы. 4). Задача. Ген цветовой слепоты и ген ночной слепоты, наследующиеся через Х хромосому, находятся на расстоянии 50 морганид друг от друга. Оба признаки рецессивны. Определите вероятность рождения детей одновременно с обеими аномалиями в семье, где жена имеет нормальное зрение, но мать ее страдала ночной слепотой, а отец – цветовой слепотой, муж же нормален в отношении обоих признаков. нормальное ночное видение - А Ночная слепота – а Нормальное цветное зрение – В Цветовая слепота – b Р ХаВХАb x XABY G ХАb, XaB; XAB,Y F1 XABXaB XABXAb XaBY XAbY Так как расстояние 50 морганид, то в 50% кроссинговера возникнут гаметы G XAB, Xab F1 XABXAB, XABXab, XABY, XabY Ответ: вероятность рождения детей в этой семье с обоими аномалиями равна 12,5% Билет№27 1).Генные мутации. Причина их возникновения. Классификация генных мутаций. Генные, или точковые, мутации связаны с изменением состава или последовательности нуклеотидов в пределах участка ДНК - гена. Нуклеотид внутри гена может быть заменен на другой или потерян, может быть вставлен лишний нуклеотид и т.д. Генные мутации могут привести к тому, что мутантный ген либо перестанет работать и тогда не образуются соответствующие и-РНК и белок, либо синтезируется белок с измененными свойствами, что приводит к изменению фенотипических признаков особи. Вследствие генных мутаций образуются новые аллели, что имеет большое эволюционное значение. В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. Если под действием мутации изменяется один нуклеотид, говорят о точковых мутациях . Точковые мутации с заменой оснований разделяют на два класса: транзиции (замена пурина на пурин или пиримидина на пиримидин) и трансверсии (замена пурина на пиримидин или наоборот). Из-за вырожденности генетического кода могут быть три генетических последствия точковых мутаций: сохранение смысла кодона ( синонимическая замена нуклеотида ), изменение смысла кодона, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи ( миссенс-мутация ) или образование бессмысленного кодона с преждевременной терминацией ( нонсенс-мутация ). В соответствии с этим получают название и мутации, приводящие к образованию бессмысленных триплетов (например амбер-мутация). Причины возникновения мутаций По причинам возникновения различают спонтанные и индуцированные мутации. Спонтанные (самопроизвольные) мутации возникают без видимых причин. Эти мутации иногда рассматривают как ошибки трех Р: процессов репликации, репарации и рекомбинации ДНК. Это означает, что процесс возникновения новых мутаций находится под генетическим контролем организма. Например, известны мутации, которые повышают или понижают частоту других мутаций; следовательно, существуют гены-мутаторы и гены-антимутаторы. В то же время, частота спонтанных мутаций зависит и от состояния клетки (организма). Например, в условиях стресса частота мутаций может повышаться. Индуцированные мутации возникают под действием мутагенов. Мутагены – это разнообразные факторы, которые повышают частоту мутаций. Впервые индуцированные мутации были получены отечественными генетиками Г.А. Надсоном и Г.С. Филипповым в 1925 г. при облучении дрожжей излучением радия. Различают несколько классов мутагенов: – Физические мутагены: ионизирующие излучения, тепловое излучение, ультрафиолетовое излучение. – Химические мутагены: аналоги азотистых оснований (например, 5-бромурацил), альдегиды, нитриты, метилирующие агенты, гидроксиламин, ионы тяжелых металлов, некоторые лекарственные препараты и средства защиты растений. – Биологические мутагены: чистая ДНК, вирусы, антивирусные вакцины. – Аутомутагены – промежуточные продукты обмена веществ (интермедиаты). Например, этиловый спирт сам по себе мутагеном не является. Однако в организме человека он окисляется до ацетальдегида, а это вещество уже является мутагеном. 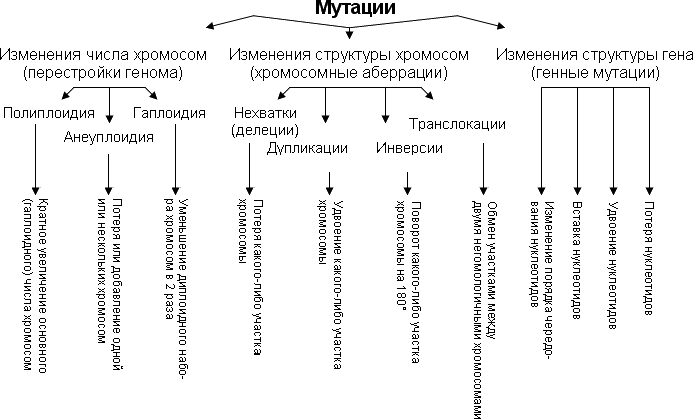 2).Основные положения хромосомной теории. Кроссинговер как механизм, определяющий нарушение сцепления генов. 1. Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален. 2. Аллельные гены занимают одинаковые локусы в гомологичных хромосомах. 3. Гены расположены в хромосоме в линейной последовательности на определенном расстоянии друг от друга. 4. Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцеплено (совместно), благодаря чему происходит сцепленное наследование. Число групп сцепления равно гаплоидному числу хромосом данного вида у (гомогаметного пола) или больше на 1 (у гетерогаметного пола). 5. Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами). Причина нарушения сцепления заключается в том, что в ходе мей-оза происходит кроссинговер и гомологичные хромосомы обмениваются своими участками. В результате гены, расположенные в одной из гомологичных хромосом, оказываются в другой хромосоме. Возникают новые сочетания признаков Кроссинговер (от англ. crossingover), перекрест, взаимный обмен участками парных хромосом, происходящий в результате разрыва и соединения в новом порядке их нитей — хроматид (рис.); приводит к перераспределению (рекомбинации) сцепленных генов. Т. о., К. — важнейший механизм, обеспечивающий комбинаторную изменчивость, а следовательно, — один из главных факторов эволюции. К., как правило, имеет место в профазе первого деления половых клеток (см. Мейоз), когда их хромосомы представлены четырьмя нитями. В месте перекреста удаётся цитологически обнаружить характерную фигуру перекрещенных хромосом — хиазму. Результат К. можно выявить по новому сочетанию сцепленных генов (если аллели гомологичных хромосом, участвовавших в К., были гетерозиготны). Этот приём, открытый американским генетиком Т. Морганом, позволил доказать линейное размещение генов в хромосоме и разработать метод установления их взаиморасположения (см. Генетические карты хромосом). В 1933 немецкий учёный К. Штерн цитологически доказал осуществление К. при обмене генами между хромосомами. Частота К. в грубом приближении зависит от линейного расстояния между генами. В случае, если на участке между двумя генами происходит сразу двойной или множественный обмен, частота перекомбинации этих генов уменьшается. Если разрывы в хромосомах, обменивающихся участками, произойдут не в строго идентичных точках, то наступит так называемый неравный К. При этом одна из хромосом получит дополнительный генетический материал, а в гомологичной хромосоме окажется его нехватка. У высших организмов обнаружен К. и в клетках тела (соматических), в этом случае он приводит к формированию мозаичных признаков. К. может захватывать обе нити молекулы ДНК или только одну; он может затронуть большой участок хромосомы с несколькими генами или часть одного гена (внутригенный К.) 3).Тип Плоские черви. Морфология, жизненный цикл, пути инвазии, локализация, патогенное действие бычьего цепня. Особенности лабораторной диагностики и профилактики тениаринхоза. ТИП ПЛОСКИЕ ЧЕРВИ (PLATHELMINTHES) Тип плоские черви включает около 7300 видов. Свободноживущие плоские черви обитают в морских и пресных водах, почве. Многие воды перешли к паразитическому образу жизни. Морфологическая характеристика типа Для плоских червей характерны следующие признаки: 1)трех-слойнойсть, т.е. развитие экто-, энто-, и мезодермы у эмбрионов; 2) наличие кожно-мускульного мешка; 3) отсутствие полости тела (пространство между органами заполнено паренхимой); 4) билатеральная симметрия; 5) форма тела, сплюснутая в дорсовентральном направлении; 6)наличие развитых систем органов: мышечной, пищеварительной, выделительной, нервной и половой. Указанные признаки-следствия ароморфозов, произошедших у предков плоских червей. Плоские черви-первичноротые животные. Тип плоские черви включает три класса: Ресничные (Turbellaria), Сосальщики (Trematodes), и Ленточные (Cestoidea). Бычий, или невооруженный цепень Taeniarhynchus saginatus - возбудитель тениаринхоза. Половозрелая форма локализуется в тонком кишечнике человека. Распространен повсеместно, встречается чаще, чем свиной цепень. Наиболее часто тениаринхоз отмечается в Закавказье, в странах СНГ, особенно часто в Средней Азии, Дагестане. Один из самых крупных гельминтов человека. Стробила лентовидной формы состоит из большого числа члеников (до 200), достигает длины 7-10 м(до 18см). По строению сходен со свиным цепнем. Отличительным признаком является отсутствие крючьев на сколексе (отсюда название невооруженный цепень). Органами фиксации являются 4 присоски. Гермафродитные проглоттиды (9х12 мм) крупнее чем у свиного цепня, квадратной формы и не имеют третьей дополнительной дольки яичника. Зрелые членики прямоугольной формы, сильно вытянуты. Замкнутая матка очень разветвлена, число ее боковых ветвей достигает 17-35 пар. Зрелые яичники способны к активному движению. Выделяются обычно по одиночке, активно выползают из анального отверстия и передвигаются по телу и белью. Несмотря на почти ежедневное отделение до 6-8 члеников и более, длина цепня не уменьшается, так как новые членики образуются в зоне роста. Срок жизни бычьего цепня до 20 лет и более. Жизненный цикл бычьего цепня типичен. Окончательный хозяин только человек, промежуточный-крупный рогатый скот. При попадании члеников или испражнений в окружающую среду происходит массовое загрязнение онкосферами почвы, травы, сена, мест содержания скота, пастбищь. С загрязненным кормом онкосферы попадают в организм крупного рогатого скота, где, осаждаясь в мышцах, превращаются в финны- цистицерки, которые могут сохранятся в теле скота до 1-2 лет. Заражение человека происходит при употреблении в пищу сырого или полусырого говяжьего мяса, содержащего финны (финнозное мясо). Финны также могут сохранять жизнеспособность в шашлыке, недостаточно проваренном мясе, в строганине из свежезамороженого мяса, в говяжьем фарше. В тонком кишечнике человека головка финны выворачиваются, прикрепляется к его стенке и через 3 месяца превращается во взрослого половозрелого паразита. Восприимчивость к териаринхозу всеобщая, но паражаемость взрослых значительно выше, чем детей, что связано с особенностями питания. Женщины поражаются чаще чем мужчины, среди последних инвазия встречается обычно в профессиональных группах, связанных с забоем животных и приготовлением пищи (рабочие боен, мясокомбинатов, повара). Поражаемость таниаринхозом высока в сельской местности, особенно в районах с развитым скотоводством. В патогенезе тениархиоза имеют значение повреждение слизистой оболочки тонкой кишки присосками гельминта, раздражение механорецепторов, токсическое воздействие на организм продуктов обмена червя, а также обеднение организма человека пищевыми веществами в результате интенсивной резорбции их гельминтом и блокирования некоторых ферментов в кишечнике человека антиферментами паразита. Длительность инвазии составляет несколько лет. Известны случаи паразитирования у человека нескольких особей бычьего цепня. Симптомы инвазии обычно появляются в хронической фазе. Считается что по патогенному действию тениаринхоз менее опасен, чем тениоз, так как не дает осложнения в виде цестицеркоза. Однако, большие размеры и почти ежедневное активное выхождение члеников из кишечника инвазированного человека дают более значительную сумму механических, рефлекторных и токсикоаллергических влияний на организм хозяина. Отмечены диспептические расстройства, схваткообразные боли в животе, поносы и общая слабость, повышение аппетита, сочетающегося с потерей веса. Снижается кислотность желудочного сока. У ослабленных лиц инвазия сопровождается неврологическими симптомами, обмороками, головокружениями, нарушением сна, судорожными припадками. Осложнения появляются крайне редко, возможно заползание проглоттид в червеобразный отросток или влагалище, атипичная локализация цепня в желчном пузыре, двенадцатиперстной кишке, поджелуджочной железе, в брюшной полости. Известны случаи обтурационной непроходимости кишечника, обусловленной множественной инвазией цепнями. Лабораторная диагностика проводится макровогельминтоскопическим методом- обнаружение в фекалиях зрелых проглоттид и подсчет кол-ва боковых ветвей матки (от 17 до 35 с каждой стороны), а также путем опроса больного (зрелые членики выделяются по одиночке, способны к активному движению) 4).Задача. Аниридия наследуется как доминантный аутосомный признак и встречается с частотой 1:10 000 (В. П. Эфроимсон). Определите генетическую структуру популяции. Используем закон Харди-Вайнберга. Из этого закона следует, что число доминантных гомозигот равно квадрату частоты доминантного гена (р2), число гомозиготных рецессивных особей равно квадрату частоты рецессивного гена (q2), число гетерозиготных генотипов равно удвоенному произведению частот обоих аллелей (2pq). Частота генов в популяции выражается формулой р +q= I. Билет 28 1). Хромосомные аберрации (хромосомныемутации, хромосомные перестройки) — тип мутаций, которые изменяют структуру хромосом. Классифицируют: делеции (утрата участка хромосомы), инверсии (изменение порядка генов участка хромосомы на обратный), дупликации (повторение участка хромосомы), транслокации (перенос участка хромосомы на другую), а также дицентрические и кольцевые хромосомы. Известны также изохромосомы, несущие два одинаковых плеча. Если перестройка изменяет структуру одной хромосомы, то такую перестройку называют внутрихромосомной (инверсии, делеции, дупликации, кольцевые хромосомы), если же двух разных, то межхромосомной (дупликации, транслокации, дицентрические хромосомы). Хромосомные перестройки подразделяют также на сбалансированные и несбалансированные. Сбалансированные перестройки (инверсии, реципрокные транслокации) не приводят к потере или добавлению генетического материала при формировании, поэтому их носители, как правило, фенотипически нормальны. Несбалансированные перестройки (делеции и дупликации) меняют дозовое соотношение генов, и, как правило, их носительство сопряжено с клиническими отклонениями от нормы. Хромосомные перестройки играют определенную роль в эволюционном процессе и видообразовании, в нарушении фертильности, в онкологических и врождённых наследственных заболеваниях человека. Хромосомные перестройки были открыты у дрозофил при помощи генетического анализа. В некоторых скрещиваниях соотношение числа потомков в разных классах сильно отличалось от ожидаемого, и это объяснили наличием перестроек в хромосомах родителей. Делеции, дупликации и транслокации обнаружил К. Бриджес в 1916, 1919 и 1923 годах, соответственно. Первую инверсию описал А. Стёртевант в 1921 году, сравнивая порядок генов в хромосоме 3 у D.melanogaster и D.simulans. Первые наблюдения хромосомных перестроек были сделаны на политенных хромосомах слюнных желез. Лишь спустя некоторое время существование перестроек было доказано цитологически на митотических хромосомах. Однако проще всего перестройки можно увидеть в политенных хромосомах у гетерозиготных особей, благодаря образованию петель и крестообразных структур. Также перестройки можно увидеть в профазе мейоза при образовании синаптонемных комплексов, где, благодаря синапсису гомологичных хромосом, также образуются петли и крестообразные структуры.[1]:1 |