Биохимия. ЭКЗАМЕНАЦИОННЫЕ ВОПРОСЫ ПО БИОЛОГИЧЕСКОЙ ХИМИИ. Экзаменационные вопросы по биологической химии для студентов лечебного, педиатрического и медикопрофилактического факультетов
 Скачать 6.22 Mb. Скачать 6.22 Mb.
|

|
Нейросклеропротеиныможно охарактеризовать как структурно-опорные белки. Основные представители этих белков – нейроколлагены, нейроэлас-тины, нейростромины и др. Они составляют примерно 8–10% от общего количества простых белковнервной ткани и локализованы в основном в белом веществе головного мозга и в периферической нервной системе. Сложные белки нервной ткани представлены нуклеопротеинами, липопротеинами, протеолипидами, фосфопротеинами,гликопротеинами и т.д. В мозговой ткани содержатся в значительном количестве еще более сложные надмолекулярные образования, такие, как липонуклеопротеины, липогликопротеины и, возможно, липогликонуклеопротеиновые комплексы. Нуклеопротеины– белки, которые принадлежат либо к дезоксирибонук-леопротеинам, либо к рибонуклеопротеинам. Часть этих белковиз мозговой ткани извлекается водой, другая часть – солевыми средами, а третья – 0,1 М раствором щелочи. Липопротеинысоставляют значительную часть водорастворимых белков мозговой ткани. Их липидный компонент – это в основномфосфоглицериды и холестерин. Протеолипиды– это белково-липидные соединения, экстрагируемые органическими растворителями из ткани мозга. Отличаются от водорастворимых липопротеинов тем, что они нерастворимы в воде, но растворимы в смеси хлороформ–метанол. Белки, освобожденные от липидов, растворимы в воде, а также (благодаря высокому содержанию гидрофобных аминокислот) в смеси хлороформ–метанол. Наибольшее количество про-теолипидов сосредоточено в миелине, в небольших количествах они входят в состав синаптических мембран и синаптических пузырьков. Фосфопротеиныв головном мозге содержатся в большем количестве, чем в других органах и тканях,– около 2% от общего количества всех сложных белков мозга. Фосфопротеины обнаружены в мембранах различных морфологических структур нервной ткани. Гликопротеиныпредставляют собой чрезвычайно гетерогенную группу белков. По количеству белка и углеводов, входящих в состав гликопро-теинов, их можно разделить на две основные группы. Первая группа – это гликопротеины, содержащие от 5 до 40% углеводови их производных; белковая часть состоит преимущественно из альбуминов и глобулинов. В гликопротеинах, составляющих вторую группу, содержится 40–85% углеводов, часто обнаруживается липидный компонент; по своему составу они могут быть отнесены к гликолипопротеинам. В нервной ткани обнаружен ряд специфических белков, в частности белок S-100 и белок 14-3-2. Белок S-100, или белок Мура, называют также кислым белком, так как он содержит большое количество остатков глутаминовой и аспарагиновой кислот. Этот белоксосредоточен в основном в нейроглии (85-90%), в нейронах его не более 10-15% от общего количества белка в головном мозге. Установлено, что концентрация белка S-100 возрастает при обучении (тренировках) животных. Пока нет оснований считать, что белокS-100 непосредственно участвует в формировании и хранении памяти. Не исключено, что его участие в этих процессах опосредованно.Белок 14-3-2 также относится к кислым белкам. В отличие от белка S-100 он локализован в основном в нейронах; в нейроглиальныхклетках его содержание невелико. Пока неясна роль белка 14-3-2 в выполнении специфических функций нервной ткани. Ферменты. В мозговой ткани содержится большое количество ферментов, катализирующих обмен углеводов, липидов и белков. До сихпор в кристаллическом виде из ЦНС млекопитающих выделены лишь некоторые ферменты, в частности ацетилхолинэстераза и креатинкиназа. Значительное количество ферментов в мозговой ткани находится в нескольких молекулярных формах (изоферменты): ЛДГ, альдолаза, креатинкиназа, гексокиназа, малатдегидрогеназа, глутаматдегидрогеназа, холинэстераза, кислая фосфатаза, моноаминоксидаза и др. Среди химических компонентов головного мозга особое место занимают липиды, высокое содержание и специфическая природа которых придают мозговой ткани характерные особенности. В группу липидов головного мозга входят фосфоглицериды, холестерин,сфингомиелины, цереброзиды, ганглиозиды и очень небольшое количество нейтрального жира. Многие липиды нервной ткани находятся в тесной взаимосвязи с белками, образуя сложные системы типа протеолипидов. В сером веществе головного мозга фосфоглицериды составляют более 60% от всех липидов, а в белом веществе – около 40%. Напротив, в белом веществе содержание холестерина, сфингомиелинов и особенно церебрози-дов больше, чем в сером веществе. В мозговой ткани имеются гликоген и глюкоза, но по сравнению с другими тканями ткань мозга бедна углеводами. Общее содержаниеглюкозы в головном мозге разных животных составляет в среднем 1–4 мкмоль на 1 г ткани, а гликогена – 2,5–4,5 мкмоль на 1 г ткани. Интересно отметить, что общее содержание гликогена в мозге эмбрионов и новорожденных животных значительно выше, чем в мозге взрослых. Например, у новорожденных мышей в отличие от взрослых особей уровень гликогена в 3 раза выше. По мере роста и дифференцировки мозга концентрация гликогена быстро снижается и остается относительно постоянной у взрослого животного. В мозговой ткани имеются также промежуточные продукты обмена углеводов: гексозо- и триозофосфаты, молочная, пировиноградная и другие кислоты . В мозговой ткани на долю адениновыхнуклеотидов приходится около 84% от всех свободных нуклеотидов. Большую часть оставшихсянуклеотидов составляют производные гуанина. В целом количество высокоэргических соединений в нервной ткани невелико. Содержание нуклеотидов и креатин-фосфата в головном мозге крыс составляет в среднем (в мкмоль на 1 г сырой массы): АТФ – 2,30–2,90; АДФ – 0,30–0,50; АМФ – 0,03–0,05; ГТФ – 0,20–0,30; ГДФ – 0,15–0,20; УТФ – 0,17–0,25; креатинфосфат – 3 , 5 0 – 4,75. Распределение основных макроэргических соединений примерно одинаково во всех отделах мозга. Содержание циклических нуклеотидов (цАМФ и цГМФ) в головном мозге значительно выше, чем во многих других тканях. Уровень цАМФв мозге в среднем 1–2, а цГМФ – до 0,2 нмоль на 1 г ткани. Для мозга характерна также высокая активность ферментов метаболизмациклических нуклеотидов. Большинство исследователей считают, что циклические нуклеотиды участвуют в синаптической передаче нервного импульса. Ионы Na+, K+, Cu2+, Fe3+, Ca2+, Mg2+и Мn2+ распределены в головном мозге относительно равномерно в сером и белом веществе. Содержание фосфатов в белом веществе выше, чем в сером. Количественное соотношение неорганических анионов и катионов в мозговой ткани свидетельствует о дефиците анионов. Расчет показывает, что для покрытия дефицита анионов потребовалось бы в 2 раза больше белков, чем их имеется в мозговой ткани. Принято считать, что остающийся дефицит анионов покрывается за счет липидов. Вполне возможно, что участие липидов в ионном балансе – одна из функций головного мозга. Миелиновоевещество – понятие морфологическое. По сути миелин – это система, образованная многократно наслаивающимисямембранами клеток нейроглии вокруг нервных отростков (в периферических нервных стволах нейроглия представлена леммоцитами, или шванновскими клетками, а в белом веществе ЦНС – астроцитами). По химическому составу миелиновое вещество является сложным белково-липидным комплексом. На долю липидов приходится до 80% плотного осадка; 90% всех липидов миелина представлено холестерином, фосфолипидами и церебро-зидами. Считают, что в липоидных слоях миелиновых оболочек молекулы различных липидов имеют строго определенное расположение. 166. Энергетический обмен в нервной ткани. Значение аэробного распада глюкозы. На долю головного мозга приходится 2–3% от массы тела. В то же время потребление кислорода головным мозгом в состоянии физического покоя достигает 20–25% от общего потребления его всем организмом, а у детей в возрасте до 4 лет мозг потребляет даже 50% кислорода, утилизируемого всем организмом. О размерах потребления головным мозгом из крови различных веществ, в том числе кислорода, можно судить по артериовенозной разнице. Установлено, что во время прохождения через мозг кровь теряет около 8 об.% кислорода. В 1 мин на 100 г мозговой тканиприходится 53–54 мл крови. Следовательно, 100 г мозга потребляет в 1 мин 3,7 мл кислорода, а весь головной мозг (1500 г) – 55,5 млкислорода. Газообмен мозга значительно выше, чем газообмен других тканей, в частности он превышает газообмен мышечной ткани почти в 20 раз. Интенсивность дыхания для различных областей головного мозга неодинакова. Например, интенсивность дыхания белого веществав 2 раза ниже, чем серого (правда, в белом веществе меньше клеток). Особенно интенсивно расходуют кислород клетки коры мозга и мозжечка. Поглощение кислорода головным мозгом значительно меньше при наркозе. Напротив, интенсивность дыхания мозга возрастает при увеличении функциональной активности. Основным субстратом дыхания мозговой ткани является глюкоза. В 1 мин 100 г ткани мозга потребляют в среднем 5 мг глюкозы. Подсчитано, что более 90% утилизируемой глюкозы в ткани мозга окисляется до СО2 и Н2О при участии цикла трикарбоновых кислот. В физиологических условиях роль пентозофосфатного пути окисления глюкозы в мозговой ткани невелика, однако этот путь окисления глюкозы присущ всем клеткам головного мозга. Образующаяся в процессе пентозофосфатного цикла восстановленная форма НАДФ (НАДФН) используется для синтеза жирных кислот и стероидов. Интересно отметить, что в расчете на всю массу головного мозга содержание глюкозы в нем составляет около 750 мг. За 1 мин тканью мозга окисляется 75 мг глюкозы. Следовательно, количество глюкозы, имеющееся в ткани головного мозга, могло бы быть достаточным лишь на 10 мин жизни человека. Данный расчет, а также величина артериовенозной разницы по глюкозе доказывают, что основным субстратом дыхания головного мозга является глюкозакрови. По-видимому, глюкоза легко диффундирует из крови в ткань головного мозга (содержание глюкозы в мозговой ткани 0,05%, а в артериальной крови – 4,44 ммоль/л, или 80 мг/100 мл). Между глюкозой и гликогеном мозговой ткани имеется тесная связь, выражающаяся в том, что при недостаточном поступлении глюкозыиз крови гликоген головного мозга является источником глюкозы, а глюкоза при ее избытке – исходным материалом для синтеза гликогена. Распад гликогена в мозговой ткани происходит путем фосфоролиза с участием системы цАМФ. Однако в целом использование гликогена в мозге по сравнению с глюкозой не играет существенной роли в энергетическом отношении, так как содержание гликогена в головном мозге невелико. Наряду с аэробным метаболизмом углеводов мозговая ткань способна к довольно интенсивному анаэробному гликолизу. Значение этого явления пока недостаточно ясно, ибо гликолиз как источник энергии ни в коей мере не может сравниться по эффективности с тканевым дыханием в головном мозге. Интенсивность обновления богатых энергией фосфорных соединений в головном мозге очень велика. Именно этим можно объяснить, что содержание АТФ и креатинфосфата в мозговой ткани характеризуется значительным постоянством. В случае прекращения доступакислорода мозг может «просуществовать» немногим более минуты за счет резерва лабильных фосфатов. Прекращение доступакислорода даже на 10–15 с нарушает энергетику нервных клеток, что в целостном организме выражается наступлением обморочного состояния. По-видимому, при кислородном голодании мозг может очень недолго получать энергию за счет процессов гликолиза. Установлено, что при инсулиновой коме содержание глюкозы в крови может снижаться до 1 ммоль/л, потребление кислорода мозгом в этих условиях не более 1,9 мл/100 г в 1 мин. В норме концентрация глюкозы в крови 3,3–5,0 ммоль/л, а мозг потребляет 3,4–3,7 млкислорода на 100 г массы в 1 мин. При инсулиновой коме нарушаются процессы окислительного фосфорилирования в мозговой ткани, снижается концентрация АТФ и происходит изменение функций мозга. Возбуждение и наркоз быстро сказываются на обмене лабильных фосфатов. В состоянии наркоза наблюдается угнетение дыхания; содержание АТФ и креатинфосфата повышено, а уровень неорганического фосфата снижен. Следовательно, сокращается потребление мозгом соединений, богатых энергией. Напротив, при раздражении интенсивность дыхания усиливается в 2–4 раза; уровень АТФ и креатинфосфата снижается, а количествонеорганического фосфата увеличивается. Эти изменения наступают независимо от того, каким образом произошло стимулирование нервных процессов, а именно путем электрического разряжения или химическим путем. Общее содержание аминокислот в ткани мозга человека в 8 раз превышает концентрацию их в крови. Аминокислотный состав мозга отличается определенной специфичностью. Так, концентрация свободной глутамино-вой кислоты в мозге выше, чем в любом другом органе млекопитающих (10 мкмоль/г). На долю глутаминовой кислоты вместе с ее амидом глу-тамином и трипептидом глутатиономприходится более 50% α-аминоазота головного мозга. В мозге содержится ряд свободных аминокислот, которые лишь в незначительных количествах обнаруживаются в других тканях млекопитающих. Это γ-аминомасляная кислота, N-ацетиласпарагиновая кислота и цистатионин. Известно, что обмен аминокислот в мозговой ткани протекает в разных направлениях. Прежде всего пул свободных аминокислотиспользуется как источник «сырья» для синтеза белков и биологически активных аминов. Одна из функций дикарбоновых аминокислот в головном мозге – связывание аммиака, освобождающегося при возбуждении нервных клеток. Поступления аминокислот в мозговую ткань и выход из нее, а также использование глюкозы крови для синтеза аминокислот нейронов и глии в разных отделах мозга различны. Эти различия в существенной мере обусловлены наличием гематоэнцефалического барьера, который следует рассматривать конкретно для каждого вещества или класса веществ. Ге-матоэнцефалический барьер не следует представлять как единое структурное образование, создающее преграду для транспорта; различие относительно скоростей поступления веществ в разные отделы мозга может быть обусловлено особенностями эпителия сосудов, базальной мембраны или расположения прилегающих отростков глиальных клеток. В условиях in vitro (в отсутствие барьера) многие аминокислотынакапливаются в клетках мозга за счет активного транспорта, в котором участвует несколько самостоятельных Na+-зависимых транспортных систем. Установлено, что белки в головном мозге находятся в состоянии активного обновления, о чем свидетельствует быстрое включение радиоактивных аминокислот в молекулы белков. Однако в разных отделах головного мозга скорость синтеза и распада белковыхмолекул неодинакова. Белки серого вещества полушарий большого мозга и белки мозжечка отличаются особенно большой скоростью обновления. В участках головного мозга, богатых проводниковыми структурами – аксонами (белое вещество головного мозга), скорость синтеза и распада белковых молекул меньше. При различных функциональных состояниях ЦНС наступают изменения в интенсивности обновления белков. Так, при действии наорганизм животных возбуждающих агентов (фармакологические средства и электрический ток) в головном мозге усиливается интенсивность обмена белков. Под влиянием наркоза скорость распада и синтеза белков снижается. Возбуждение нервной системы сопровождается повышением содержания аммиака в нервной ткани. Это явление наблюдается как при раздражении периферических нервов, так и при раздражении мозга. Считают, что образование аммиака при возбуждении в первую очередь происходит за счет дезаминирования АМФ. Аммиак – очень ядовитое вещество, особенно для нервной системы. Особую роль в устранении аммиака играет глутаминовая кислота. Она способна связывать аммиак с образованием глутамина – безвредного для нервной ткани вещества. Данная реакция амидирования протекает при участии фермента глутаминсинтетазы и требует затраты энергии АТФ. Непосредственный источникглутаминовой кислоты в мозговой ткани – путь восстановительного аминирования α-кетоглутаровой кислоты;  Образование глутаминовой кислоты из α-кетоглутаровой и аммиака является важным механизмом нейтрализации аммиака в тканимозга, где путь устранения аммиака за счет синтеза мочевины не играет существенной роли. Кроме того, глутаминовая кислота в нервной ткани может декарбокси-лироваться с образованием ГАМК: 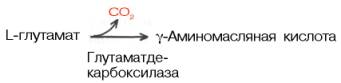 ГAMК в наибольшем количестве содержится в сером веществе головного мозга. В спинном мозге и периферических нервах ее значительно меньше. |