Экзаменационные вопросыответы на экзамен по биохимии для педиатрического факультета 2012 года
 Скачать 4.91 Mb. Скачать 4.91 Mb.
|

|
28. Механизмы репликации ДНК (матричный принцип, полуконсервативный способ). Условия, необходимые для репликации. Этапы репликации Механизмы РЕПЛИКАЦИЯ - процесс самоудвоения ДНК. В основе механизма репликации лежит принцип комплиментарности. К механизму репликации относится матричный биосинтез. Репликация ДНК идёт полуконсервативным способом: на каждой материнской полинуклеотидной цепи синтезируется дочерняя цепь. Условия необходимые для репликации: 1. Матрица - нити ДНК. Расщепление нити называется РЕПЛИКАТИВНАЯ ВИЛКА 2. Субстрат. Пластическим материалом являются ДЕЗОКСИНУКЛЕОТИДТРИФОСФАТЫ: дАТФ, дГТФ, дЦТФ, дТТФ. 3. Ионы магния. Репликативный комплекс ферментов: A) ДНК -раскручивающие белки:
B) Белки, препятствующие соединению нитей ДНК (SSB -белки) C) ДНК-ПОЛИМЕРАЗА (катализирует образование фосфодиэфирных связей). ДНК- ПОЛИМЕРАЗА только удлиняет уже существующую нить, но не может соединить два свободных НУКЛЕОТИДА. D) ПРАЙМАЗА (катализирует образование «затравки» к синтезу). Е)ДНК-ЛИГАЗА. 5. ПРАЙМЕРЫ - «затравка» для репликации. Это короткий фрагмент из рИБОНУКЛЕОТИДТРИФОСФАТОВ (2 - 10).. Основные этапы репликации. I. ИНИЦИАЦИЯ репликации. Происходит под влиянием внешних стимулов (факторов роста). Белки соединяются с рецепторами на плазматической мембране и вызывают репликацию в синтетическую фазу клеточного цикла. Смысл инициации заключается в присоединении в точку репликации DNА-А, стимулирующего расхождение двойной спирали. В этом принимает участие и ХЕЛИКАЗА. Действуют ферменты (ТОПОИЗОМЕРАЗЫ), вызывающие раскручивание сверх спирали. SSВ-белки препятствуют соединению дочерних цепей. Образуется РЕПЛИКАТИВНАЯ ВИЛКА. 2. Образование дочерних нитей. Этому предшествует образование ПРАЙМЕРОВ с помощью ПРАЙМАЗЫ. Действует ДНК-ПОЛИМЕРАЗА и образуется дочерняя нить ДНК. Этот процесс происходит по принципу комплиментарности, и синтез идёт от 5* к 3* концу синтезируемой нити. 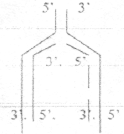 На одной из материнских нитей будет строиться непрерывная цепь, а на противоположной нити – фрагменты ОКАЗАКИ. 3. Удаление ПРАЙМЕРОВ с помощью ЭКЗОНУКЛЕАЗЫ, 4. Соединение коротких фрагментов с помощью ДНК-ЛИГАЗЫ. 28. Биосинтез РНК (транскрипция). Условия и этапы транскрипции. Процессинг РНК. Альтернативный сплайсинг Транскрипция - передача информации с ДНК на РНК (биосинтез РНК). Транскрипции подвергаются только определённые части молекулы ДНК. Эта часть называется ТРАНСКРИПТОНОМ. ДНК эукариот прерывистая: участки, несущие информацию (ЭКЗОНЫ), чередуются с участками, не несущими информацию (ИНТРОНЫ). В ДНК с 5'-конца выделяют ПРОМОТОРНУЮ область - место присоединения РНК-ПОЛИМЕРАЗЫ. С 3'-конца - ТЕРМИНАТОРНАЯ зона. Эти области не транскрибируются. УСЛОВИЯ ТРАНСКРИПЦИИ. 1. Матрица - 1 нить ДНК. Образуется транскрипционный глазок. 2. Структурные компоненты - РИБОНУКЛЕОЗИД-3-ФОСФАТЫ (АТФ, ГТФ, ЦТФ, УТФ). 3. ДНК-зависимая РНК-ПОЛИМЕРАЗА. ОСНОВНЫЕ ЭТАПЫ ТРАНСКРИПЦИИ. 1. ИНИЦИАЦИЯ. Заключается в присоединении РНК-ПОЛИМЕРАЗЫ к ПРОМОТОРУ, что приводит к расхождению нитей ДНК. Импульсом к присоединению РНК-ПОЛИМЕРАЗЫ является присоединение ТВР-белка к TATA-боксу. 2. ЭЛОНГАЦИЯ (удлинение). Соединение РИБОНУКЛЕОЗИДМОНОНУКЛЕОТИДОВ и образование фосфодиэфирных связей между НУКЛЕОТИДАМИ с помощью РНК-ПОЛИМЕРАЗЫ, которая передвигается вдоль нити ДНК. Присоединение НУКЛЕТИДОВ идет в соответствии с принципом комплиментарности, только будут РИБОНУКЛЕОТИДЫ и - УМФ. 3. ТЕРМИНАЦИЯ (окончание).Заключается в том, что со стороны 3'-конца образованной РНК присоединяется множество (до 200 - 300) АДЕНИЛОВЫХ НУКЛЕОТИДОВ - поли А. Образуется точная копия гена. АДЕНИЛОВЫЕ НУКЛЕОТИДЫ защищают 3'-конец от действия ЭКЗОНУКЛЕАЗ. С 5'-конца образуется защита, так называемый «САР» (чаще всего УДФ). Эта образовавшаяся копия гена называется ТРАНСКРИПТ. 4. ПРОЦЕССИНГ (созревание).
29. Биосинтез белка. Этапы трансляции и их характеристика. Белковые факторы биосинтеза белка. Энергетическое обеспечение биосинтеза белка. Трансляция - это перевод генетического текста М-РНК в последовательность аминокислот в белке. Характеристика белоксинтезирующего аппарата клетки. 1. МРНК - источник информации. Несоответствие числа транспортных РНК и числа кодонов (61) снимается за счёт возможности узнавания одной транспортной РНК нескольких кодонов данной аминокислоты. Транспортная РНК выполняет функцию АДАПТОРА между МРНК и белком. Фермент синтеза комплекса транспортной РНК с аминокислотой - АМИНОАЦИЛ-ТРНК-СИНТЕТАЗА (требует энергии АТФ). Белковые факторы: факторы ИНИЦИАЦИИ (ФИ) начала трансляции, факторы ЭЛОНГАЦИИ (ФЭ) - продолжатели, факторы ВЫСВОБОЖДЕНИЯ (R-факторы). Ионы магния, как КОФАКТОРЫ. АТФ, ГТФ - поддержка энергией. Этапы трансляции: 1.РЕКОГНИЦИЯ (распознавание) - узнавание между аминокислотами и их транспортной РНК. 2.АК + ТРНК аминоацил-т-РНК-синтетаза АК-ТРНК АТФАМФ + ФФ МЕТИОНИЛ-ТРНК РЕКОГНИЦИЯ происходит столько раз, сколько аминокислот входит в состав белка. 3. ИНИЦИАЦИЯ - начало процесса трансляции. На этом этапе РИБОСОМА взаимодействует с МРНК и находит стартовый кодон. Малая её единица взаимодействует с МЕТИОНИЛ-ТРНК и образует инициирующий комплекс, способный распознавать стартовый кодон. Этому предшествует разделение РИБОСОМЫ с помощью ФИ-3. Образование инициирующего комплекса происходит с помощью ФИ-2. Затем инициирующий комплекс присоединяется к МРНК с 5'-конца. Узнаванию 5'-конца способствуют САР и САР-связывающие белки. 4. ЭЛОНГАЦИЯ (продолжение) протекает циклически в виде последовательной смены трёх фаз: - присоединение следующей АМИНОАЦИЛ-ТРНК в соответствии со смыслом следующего кодона. Для процесса требуется энергия ГТФ и ФЭ-1 (проникновение в РИБОСОМУ). - ПЕПТИЗАЦИЯ. ПЕПТИДИЛТРАНСФЕРАЗА образует ПЕПТИДНУЮ связь между двумя аминокислотными остатками и одновременно разрушает сложноэфирную связь между первой аминокислотой и её ТРНК. В результате идёт образование растущего ПЕПТИДА в А-участке и высвобождение первой ТРНК - ТРАНСЛОКАЦИЯ (перемещение). При этом происходит перемещение РИБОСОМЫ на один кодон в направлении 3'-конца. При этом все остальные компоненты (МРНК, ТРНК) остаются на месте. Для процесса требуется энергия ГТФ и белковый ФЭ-2. 5. ТЕРМИНАЦИЯ (прекращение). Стоп -кодон распознаётся R-факторами (РЕЛИЗИНГ). Эти факторы высвобождают из связи компоненты белоксинтезирующего комплекса: РИБОСОМУ, МРНК, ПОЛИПЕПТИД. Помогает ПЕПТИДИЛТРАНСФЕРАЗА, отщепляющий транспортную РНК от образованного ПОЛИПЕПТИДА. Для ТЕРМИНАЦИИ нужна энергия АТФ. РИБОСОМА может повторно использоваться в трансляции. Матричная РНК или повторно используется в трансляции, или гидролизуется. ПОЛИПЕПТИД вступает в этап ПРОЦЕССИНГА белка. 6. ПРОЦЕССИНГ белка (созревание) совокупность химических модификаций ПОЛИПЕПТИДА, заканчивающихся формированием зрелой белковой молекулы. ПРОЦЕССИНГ белка может быть: КОНТРАНСЛЯЦИОННЫЙ (во время трансляции) ПОСТТРАНСЛЯЦИОННЫЙ. 31. Строение оперона. Регуляция биосинтеза белка у прокариотов. Функционирование лактозного и гистидиновых оперонов. Регуляция биосинтеза белка у ПРОКАРИОТ: 1. Регуляция происходит только на уровне транскрипции. Первичные транскрипты генов у них транслируются до завершения транскрипции. 2. Неоднородность ГЕНОМОВ. В геноме есть структурные гены и есть регуляторные области, которые могут включать регуляторные элементы и регуляторные гены. Структурные гены кодируют синтез структурных и функциональных белков. Регуляторные элементы не кодируют синтез белков вообще, но влияют на процесс транскрипции. Регуляторными элементами являются: - ПРОМОТОР - место прикрепления к ДНК РНК-ПОЛИМЕР АЗЫ, ОПЕРАТОР - место взаимодействия регуляторных белков с ДНК. Регуляторные гены кодируют синтез регуляторных белков. К ним относится белок -РЕПРЕССОР, который Может блокировать считывание информации, связываясь с оператором. Фрагмент ДНК, подверженный транскрипции называется ОПЕРОН (ПРОМОТОР, ОПЕРАТОР, структурный ген). За пределами ОПЕРОНА находятся гены-регуляторы, кодирующие синтез белка - РЕПРЕССОРА. 3. Регуляция биосинтеза белков у ПРОКАРИОТ протекает альтернативно путём репрессии и индукции. ПРИМЕР: ЛАКТОЗНЫЙ ОПЕРОН. В микробной клетке лактоза с помощью лактазы расщепляется до галактозы и глюкозы. Лактозный ОПЕРОН регулирует синтез лактазы. Если в среде присутствует лактоза, то БЕЛОК-РЕПРЕССОР вытесняется из связи с оператором и гены лактазы транскрибируются. Лактоза выступает индуктором. Гистидиновый оперон. В отсутствие гистидина белок-репрессор не имеет сродства к оператору, РНК-полимераза присоединяется к промотору, и происходит транскрипция 10 структурных генов, кодирующих строение ферментов, участвующих в синтезе гистидина; в присутствии гистидина в среде комплекс белка-репрессора с Гис, связывается с оператором, препятствует присоединению РНК-полимеразы к промотору и останавливает транскрипцию. 32. Особенности и уровни регуляции биосинтеза белка у эукариотов. Амплификация генов, энхансерные и сайленсерные элементы. Регуляция биосинтеза белка у ЭУКАРИОТ происходит на всех уровнях матричных биосинтезов. 1. На уровне транскрипции - групповая репрессия гистонами. У человека 90% ДНК репрессировано. 2. Амплификация генов - повышение числа копий гена в геноме (повышается площадь транскрипции). 3. Регуляция транскрипции сигналами-регуляторами (усилителями и душителями). Для сигналов усилителей принят термин ЭНХАЙСЕР. Они не кодируют синтез белка, чрезвычайно эффективны, может наблюдаться 200-кратное усиление транскрипции. Действие не специфично (одновременно может влиять на группу генов). Значительно удалён от ПРОМОТОРА гена-мишени. Подвержен влиянию регуляторных факторов (гормонов). Сигналы-душители называются САЙЛЕНСЕРЫ - угнетатели транскрипции. Особенности действия подобны ЭНХАЙСЕРАМ, только действие противоположное. 4. Регуляция на уровне процессинга МРНК разрешение или запрещение процессинга дифференциальный процессинг включает альтернативный СПЛАЙСИНГ - сборка РНК из разных экзонов, и редактирование МРНК - замена одного из нуклеотидов с изменением генетической информации, приводящее к образованию изменённых белков 5. На уровне стабильности и активности МРНК. МРНК в клетке образует комплекс с белками, который называется ИНФОРМОСОМА. В их составе МРНК не разрушается ферментами, сохранения в активном, стабильном состоянии. При необходимости она высвобождается из комплекса и транслируется. Процесс образования и распада ИНФОРМОСОМ регулируется гормонами. С одной молекулы РНК транслируется большое количество белков. 6. Регуляция на уровне трансляции: Тотальная регуляция может быть в виде тотальной репрессии или индукции за счёт изменения концентрации белковых факторов трансляции Избирательная дискриминация. Определённые виды МРНК избирательно не транслируются. С них не синтезируется белок. Трансляция с альтернативных стартовых участков. Т.о. образом может происходить выбор исходной стартовой точки трансляции. 33. Основные углеводы организма человека, их строение и классификация, биологическая роль. Углеводы - это ПОЛИОКСИКАРБОНИЛЬНЫЕ соединения и их производные. МОНОСАХАРИДЫ: ТРИОЗЫ (ГЛИЦЕРАЛЬДЕГИД, ДИОКСИАЦЕТОН), ТЕТРОЗЫ (ЭРИТРУЛОЗА), ПЕНТОЗЫ (РИБОЗА, ДЕЗОКСИРИБОЗА, КСИЛУЛОЗА), ГЕКСОЗЫ (ГЛЮКОЗА, ГАЛАКТОЗА, ФРУКТОЗА). ОЛИГОСАХАРИДЫ: МАЛЬТОЗА, ЛАКТОЗА, САХАРОЗА. ПОЛИСАХАРИДЫ делятся на ГОМОПОЛИСАХАРИДЫ и ГЕТЕРОПОЛИСАХАРИДЫ. ГОМОПОЛИСАХАРИДЫ - КРАХМАЛ, ГЛИКОГЕН. ГЕТЕРОПОЛИСАХАРИДЫ ХОНРОЭТИНСЕРНАЯ К-ТА, ГИАЛУРОНОВАЯ К-ТА, НЕЙРАМИНОВАЯ К-ТА, ГЕПАРИН. БИОЛОГИЧЕСКАЯ РОЛЬ УВ. 1. ЭНЕРГЕТИЧЕСКАЯ. При окислении 1 гр. УВ до конечных продуктов (СО2 и Н2О) выделяется 4,1-ккал-60-70 % всей калорийности пищи. 2. Структурная. УВ используется как строительный материал для образования структурных компонентов клеток (ГЛИКОЛИПИДЫ, ГЛИКОПРОТЕИНЫ, ГЕТЕРОПОЛИСАХАРИДЫ межклеточного вещества). 3. Резервная. УВ в виде гликогена могут откладываться в запас. 4. Защитная. ГЛИКОПРОТЕИНЫ принимают участие в образовании антител. ГИАЛУРОНОВАЯ К-ТА препятствует проникновению чужеродных веществ. ГЕТЕРОПОЛИСАХАРИДЫ участвуют в образовании слизи слизистых оболочек дыхательных путей, ЖКТ. 5. Регуляторная. Некоторые гормоны являются ГЛИКОПРОТЕИНАМИ (ТИРИОГЛОБУЛИН) 6. Участвуют в процессах распознавания клеток (СИАЛОВАЯ и НЕЙРОЛИНОВАЯ К-ТЫ). 7. Входя в состав оболочек эритроцитов, определяют группы крови. 8. Участвуют в процессах свёртывания крови, входя в состав ФИБРИНОГЕНА и ПРОТРОМБИНА. Препятствуют свёртыванию крови, входя в состав ГЕПАРИНА. 54. Роль углеводов в питании. Переваривание и всасывание углеводов в органах пищеварительной системы. Написать реакции. Непереносимость дисахаридов. Основными УВ для организма человека являются УВ пищи: крахмал, сахароза, лактоза. Поступивший с пищей крахмал в ротовой полости будет подвергаться гидролизу под действием альфа -АМИЛАЗЫ слюны. Она расщепляет альфа (1,4)-ГЛИКОЗИДНЫЕ связи. Поскольку пища в ротовой полости задерживается недолго, то крахмал здесь переваривается лишь частично. Гидролиз крахмала завершается образованием АМИЛОДЕКСТРИНОВ. Далее пища поступает в желудок. В желудке действие альфа -АМИЛАЗЫ прекращается. Однако в более глубоких слоях действие фермента, крахмал проходит следующую стадию гидролиза- ЭРИТРОДЕКСТРИНОВ. Основным местом переваривания крахмала служит тонкий отдел кишечника. В 12 п.к. открываются протоки ПЖЖ. Под действием фермента в её секрете будет идти гидролиз крахмала. Выделяющийся панкреатический сок содержит БИКАРБОНАТЫ, которые нейтрализуют желудочное кислое содержимое. Альфа -АМИЛАЗА завершает разрыв внутренних альфа(1,4)-ГЛИКОЗИДНЫХ связей с образованием ДИСАХАРИДОВ (МАЛЬТОЗ). Образованная МАЛЬТОЗА - быстро гидролизуется под действием МАЛЬТАЗ. Из тех же остатков, которые в молекулах крахмала были соединены альфа(1,6)-ГЛИКОЗИДНЫМИ связями, образуются ДИСАХАРИДЫ -ИЗОМАЛЬТОЗЫ. Они будут гидролизоваться ИЗОМАЛЬТАЗАМИ. ФОСФОРИЛИРОВАНИЕ в присутствии АТФ и фермента -ГЕКСОКИНАЗЫ. Глюкоза превращается в глюкозо-6-фосфат. Теперь он будет использоваться в АНАБОЛИЧЕСКИХ и КАТАБОЛИЧЕСКИХ реакциях. Глюкоза из клетки может выйти, если в реакции гидролиза при участии глюкозо-6-фосфатазы освободится от остатка фосфорной кислоты. Этот фермент находится в печени, почках, эпителии кишечника. Проникновение глюкозы в клетки этих органов и тканей необратимы. Процесс биосинтеза: 1) Сахароза глюкоза 2) 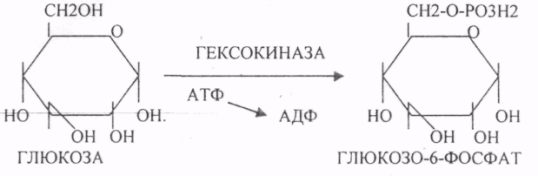 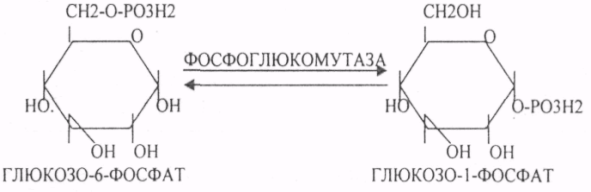 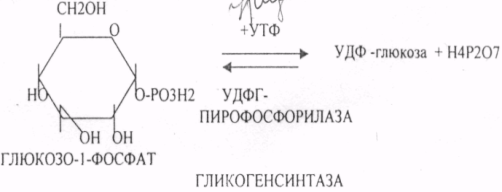 3) ГЛИКОГЕНСИНТАЗА - ТРАНСФЕРАЗА, которая переносит остатки глюкозы, входящие в УДФ- глюкозу, на ГЛИКОЗИДНУЮ связь остаточного в клетке гликогена. Образование альфа(1,6)-ГЛИКОЗИДНЫХ связей в точках ветвления гликогена катализирует ГЛИКОГЕН-ВЕТВЯЩИЙ фермент. На каждую молекулу глюкозы, включающуюся в структуру гликогена, расходуется 2 молекулы АТФ. Гликоген в клетках накапливается во время пищеварения и рассматривается как резервная форма глюкозы, которая используется клетками в промежутках между приёмами пищи. 34. Катаболизм глюкозы в анаэробных условиях. Химизм процесса, биологическая роль. АНАЭРОБНЫЙ ГЛИКОЛИЗ протекает в цитоплазме клеток. Окисление глюкозы или глюкозного остатка гликогена всегда завершается образованием конечного продукта этого процесса- лактата. Окисление глюкозы и глюкозного остатка гликогена в тканях отличается начальных стадиях превращения, до образования глюкозо-6-фосфата. Дальнейшее окисление с этого этапа в тканях как в АНА-, так и в АЭРОБНЫХ условиях полностью совпадает до стадии образования ПИРУВАТА. Процесс АНАЭРОБНОГО ГЛИКОЛИЗА сложный и многоступенчатый: -первая стадия заканчивается образованием из ГЕКСОЗЫ двух ТРИОЗ -ДИОКСИАЦЕТОНФОСФАТА и ГЛИЦЕРАЛЬДЕГИД-3-ФОСФАТА. -Вторая стадия - ГЛИКОЛИТИЧЕСКОЙ ОКСИДОРЕДУКЦИИ. Она сопряжена с образованием АТФ за счёт реакций СУБСТРАТНОГО ФОСФОРИЛИРОВАНИЯ, окислением ГЛИЦРАЛЬДЕГИД -3-ФОСФАТА, восстановлением ПИРУВАТА до ЛАКТАТА. 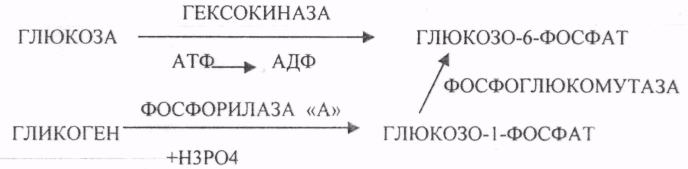 .В процессе превращения глюкозы было израсходовано 2 молекулы АТФ (ГЕКСОКИНАЗНАЯ и ФОСФОФРУКТОКИНАЗНАЯ реакции). С этапа образования ТРИОЗ идёт одновременное их окисление. В результате этих реакций образуется энергия в виде АТФ за счёт реакций СУБСТРАТНОГО ФОСФОРИЛИРОВАНИЯ (ГЛИЦЕРАТКИНАЗНАЯ и ПИРУВАТКИНАЗНАЯ). 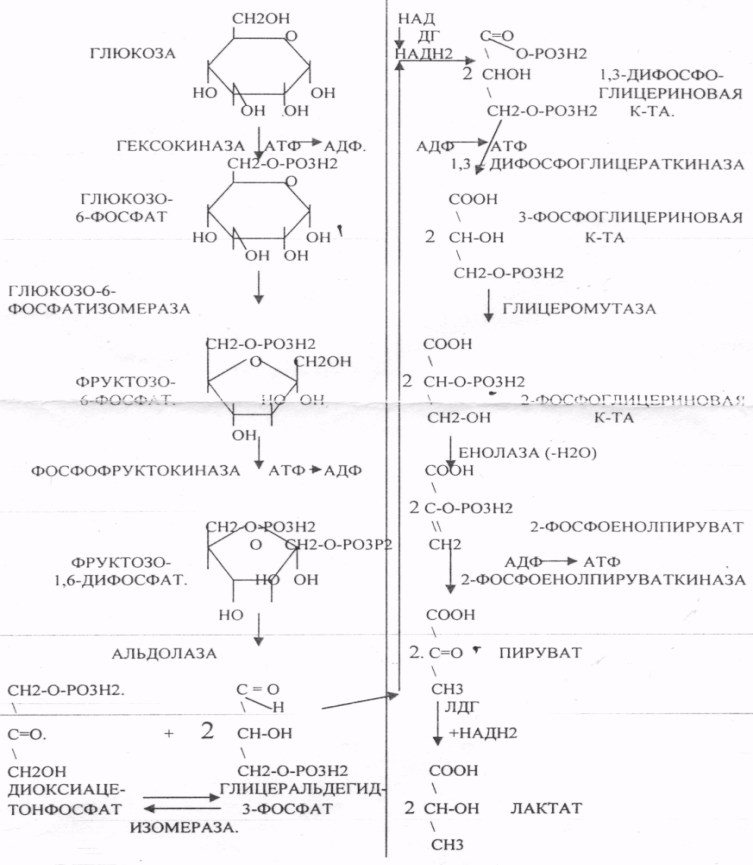 На этапе ГЛИКОЛИТИЧЕСКОЙ ОКСИДОРЕДУКЦИИ идёт окисление ГЛИЦЕРАЛЬДЕГИД-3-ФОСФАТА в присутствии НЗРО4 и НАД- зависимой ДГ. ПВК, принимая НАДН2, восстанавливается с образованием ЛАКТАТА. |