ШПОРЫ БХ 2 ИТОГОВАЯ. Эндергонические процессы, протекающие с увеличением свободной энергии. Катаболические превращения
 Скачать 2.83 Mb. Скачать 2.83 Mb.
|

|
Характеристика гормонов, регулирующих обмен гликогена Первичным сигналом для синтеза и секреции инсулина и глюкагона является изменение уровня глюкозы в крови. В норме концентрация глюкозы в крови соответствует 3,3-5,5 ммоль/л (60- 100 мг/дл). Инсулин - белковый гормон, синтезируется и секретируется в кровь р-клетками островков Лангерханса поджелудочной железы, β-клетки чувствительны к изменениям содержания глюкозы в крови и секретируют инсулин в ответ на повышение её содержания после приёма пищи. Транспортный белок (ГЛЮТ-2), обеспечивающий поступление глюкозы в β-клетки, отличается низким сродством к ней. Следовательно, этот белок транспортирует глюкозу в клетку поджелудочной железы лишь после того, как её содержание в крови будет выше нормального уровня (более 5,5 ммоль/л). В β-клетках глюкоза фосфорилируется глюкокиназой, имеющей также высокую Кm для глюкозы - 12 ммоль/л. Скорость фосфорилирования глюкозы глюкокиназой в β-клетках прямо пропорциональна её концентрации в крови. Синтез инсулина регулируется глюкозой. Глюкоза (или её метаболиты), по-видимому, непосредственно участвуют в регуляции экспрессии гена инсулина. Секреция инсулина и глюкагона также регулируется глюкозой, которая стимулирует секрецию инсулина из β-клеток и подавляет секрецию глюкагона из α-клеток. Кроме того, сам инсулин снижает секрецию глюкагона. Глюкагон - "гормон голода", вырабатываемый α-клетками поджелудочной железы в ответ на снижение уровня глюкозы в крови. По химической природе глюкагон - пептид. Адреналин выделяется из клеток мозгового вещества надпочечников в ответ на сишалы нервной системы, идущие из мозга при возникновении экстремальных ситуаций (например, бегство или борьба), требующих внезапной мышечной деятельности. Адреналин является сигналом "тревоги". Он должен мгновенно обеспечить мышцы и мозг источником энергии. Вопрос 30. наследственные нарушения обмена моносахаридов и дисахаридов. Нарушения обмена моно- и дисахаридов Фруктоземия Этиология и патогенез. Заболевание обусловлено врожденным отсутствием ферментов фруктозофосфатальдолазы и фруктозодифосфатальдолазы. Избыточное накопление фруктозофосфата нарушает гликогенолиз, что приводит к гипогликемии. В печени имеется недостаточное количество фермента фруктозо-1-фосфат-альдолазы, в результате продукты обмена (фруктозо-1-фосфат) накапливаются в организме (печени, почках, слизистых оболочках кишечника) и оказывают повреждающее действие. Морфологически в печени выявляются жировая инфильтрация, умеренный перилобулярный фиброз. Клиническая картина. Симптомы возникают при введении в рацион сладкой пищи развиваются диспептические явления и состояния острой гипогликемии, В тяжелых случаях может развиться гипогликемическая кома с потерей сознания и судорогами. Характерно, что гипогликемия возникает после приема пищи. Постоянным признаком является гепатомегалия Лечение. Диета, лишенная фруктозы, главными источниками которой считаются мед, сахарный тростник и свекла, фрукты, разрешается употреблять, животные жиры Галактоземия Этиология и патогенез. Наследственная энзимопатия. Наследуется по рецессивному типу. В основе галактоземии (рис. 1) лежит нарушение обмена галактозы в связи с отсутствием фермента галактозофосфат-уридилтрансферазы. В результате в крови накапливаются в больших концентрациях галактоза и галактозофосфат. Происходит нарушение процесса ферментативного превращения галактозы в глюкозу с накоплением галактозы и продуктов ее обмена в клетках, что оказывает повреждающее действие на функции печени, головного мозга, хрусталика глаза, почек. Клиническая картина. Клинические признаки заболевания возникают рано — через 1–2 недели после рождения ребенка. Пропадает аппетит, появляются вялость, рвота, понос. Наблюдается дефицит массы тела. Постепенно развивается гепато-, спленомегалия, появляется стойкая гипербилирубинемия, преимущест-венно за счет прямого билирубина. Часто отмечается катаракта, ведущая к слепоте. Могут быть симптомы, свидетельствующие о поражении почек (протеинурия, гипераминоацидурия), центральной нервной системы (задержка психофизического развития). После чайно-водной паузы состояние улучшается, но введение молока обусловливает рецидив нарушений со стороны желудочно-кишечного тракта. При несвоевременной диагностике заболевание прогрессирует, что приводит к тяжелым последствиям или летальному исходу. Диагностика: 1. Определение концентрации га-лактозо-1-фосфата в эритроцитах (повышена). 2. Исследование активности галак-тозо-1-фосфат-уридилтрансферазы в эритроцитах. 3. Повышение уровня галактозы в крови и моче (методом хроматографии). 4. Микробиологический тест Гатри. Лечение. Диетотерапия является единственным методом лечения. Для вскармливания ребенка используют смеси, лишенные лактозы. Из питания детей более старшего возраста исключают цельномолочные продукты. Вопрос 31 Липиды. Общая характеристика. Биологическая роль. Липи́ды - широкая группа органических соединений, включающая жирные кислоты, а также их производные, как по радикалу, так и по карбоксильной группе. Классификация липидов, как и других соединений биологической природы, — весьма спорный и проблематичный процесс. Предлагаемая ниже классификация, хоть и широко распространена в липидологии, является далеко не единственной. Она основывается, прежде всего, на структурных и биосинтетических особенностях разных групп липидов. Простые липиды Жирные кислоты, Жирные альдегиды, Жирные спирты, Предельные углеводороды с длинной алифатической цепочкой, Сфингозиновые основания, Воски, Сложные липиды Полярные (Фосфолипиды, Гликолипиды, Фосфогликолипиды, Сфинголипиды,Мышьяколипиды); Нейтральные; Ацилглицериды, Триглицериды (Жиры); Диглицериды; Моноглицериды. Церамиды, Эфиры стеринов, N-ацетилэтаноламиды, Оксилипиды, Оксилипиды липоксигеназного пути, Оксилипиды циклооксигеназного пути Биологические функции
Многие жиры, в первую очередь триглицериды, используются организмом как источник энергии. При полном окислении 1 г жира выделяется около 9 ккал энергии, примерно вдвое больше, чем при окислении 1 г углеводов (4.1 ккал). Жировые отложения используются в качестве запасных источников питательных веществ, прежде всего животными, которые вынуждены носить свои запасы на себе. Растения чаще запасают углеводы, однако в семенах многих растений высоко содержание жиров.
Жир — хороший теплоизолятор, поэтому у многих теплокровных животных он откладывается в подкожной жировой ткани, уменьшая потери тепла. у животных, обитающих в условиях жаркого климата (верблюды, тушканчики) жировые запасы откладываются на изолированных участках тела (в горбах у верблюда, в хвосте у жирнохвостых тушканчиков), в качестве резервных запасов воды, так как вода — один из продуктов окисления жиров.
Фосфолипиды составляют основу бислоя клеточных мембран, холестерин — регулятор текучести мембран. Воски образуют кутикулу на поверхности надземных органов (листьев и молодых побегов) растений. Их также производят многие насекомые пчёлы строят из них соты.
Толстый слой жира защищает внутренние органы многих животных от повреждений при ударах. Вопрос 32.классификация липидов. Характеристика отдельных групп.
Вопрос 33. высшие жирные кислоты, предельные и непредельные. Особенности строения вжк животного происхождения. Способы обозначения числа атомов углерода, положение и кол-во двойных связей. Биороль. Полиеновые ж к. Жирные кислоты - структурные компоненты различных липидов. В составе триацилгли-церолов жирные кислоты выполняют функцию депонирования энергии, так как их радикалы содержат богатые энергией СН2-группы. При окислении СН-связей энергии выделяется больше, чем при окислении углеводов, в которых атомы углерода уже частично окислены (-НСОН-). В составе фосфолипидов и сфинго-липидов жирные кислоты образуют внутренний гидрофобный слой мембран, определяя его свойства. Жиры и фосфолипиды организма при нормальной температуре тела имеют жидкую консистенцию, так как количество ненасыщенных жирных кислот преобладает над насыщенными. В фосфолипидах мембран ненасыщенных кислот может быть до 80-85%, а в составе жиров подкожного жира - до 60%. В свободном, неэтерифицированном состоянии жирные кислоты в организме содержатся в небольшом количестве, например в крови, где они транспортируются в комплексе с белком альбумином. Жирные кислоты липидов человека представляют собой углеводородную неразветвлённую цепь, на одном конце которой находится карбоксильная группа, а на другом - метальная группа (ω-углеродный атом). Большинство жирных кислот в организме содержат чётное число атомов углерода - от 16 до 20). Жирные кислоты, не содержащие двойных связей, называют насыщенными. Основной насыщенной жирной кислотой в липидах человека является пальмитиновая (до 30-35%). Жирные кислоты, содержащие двойные связи, называют ненасыщенными. Ненасыщенные жирные кислоты представлены моноеновыми (с одной двойной связью) и полиеновыми (с двумя и большим числом двойных связей). Если в составе жирной кислоты содержатся две и более двойных связей, то они располагаются через СН2-группу. Имеется несколько способов изображения структуры жирных кислот. При обозначении жирной кислоты цифровым символом общее количество атомов углерода представлено цифрой до двоеточия, после двоеточия указывают число двойных связей. Позицию двойной связи обозначают знаком Δ, после которого указывают номер атома углерода, ближайшего к карбоксилу, у которого находится двойная связь. Например, С18:1Δ9 означает, что жирная кислота содержит 18 атомов углерода и одну двойную связь у 9-го атома углерода, считая от углеродного атома карбоксильной группы. Позиция двойной связи может быть указана и другим способом - по расположению первой двойной связи, считая от метального со-атома углерода жирной кислоты. Например, линоле-вая кислота может быть обозначена как С18:2Δ9,12 или С18:2ω-6. По положению первой двойной связи от метального углерода полиеновые жирные кислоты делят на семейства ω-3 и ω-6. Двойные связи в жирных кислотах в организме человека имеют цис-конфигурацию. Это означает, что ацильные фрагменты находятся по одну сторону двойной связи. Цис-конфигурация двойной связи делает алифатическую цепь жирной кислоты изогнутой, что нарушает упорядоченное расположение насыщенных радикалов жирных кислот в фосфолипидах мембран) и снижает температуру плавления. Чем больше двойных связей в жирных кислотах липидов, тем ниже температура их плавления. Жирные кислоты с транс-конфигурацией двойной связи могут поступать в организм с пищей, например в составе маргарина. В этих кислотах отсутствует излом, характерный для цис-связи, поэтому жиры, содержащие такие ненасыщенные кислоты, имеют более высокую температуру плавления, т.е. более твёрдые по консистенции. Большинство жирных кислот синтезируется в организме человека, однако полиеновые кислоты (линолевая и α-линоленовая) не синтезируются и должны поступать с пищей. Эти жирные кислоты называют незаменимыми, или эссенциальными. Основные источники полиеновых жирных кислот для человека - жидкие растительные масла и рыбий жир, в котором содержится много кислот семейства ω-3 Жиры, содержащие преимущественно насыщенные кислоты, являются твёрдыми (говяжий, бараний жиры), а содержащие большое количество ненасыщенных кислот - жидкими. Жидкие жиры или масла обычно имеют растительное происхождение Из животных пищевых жиров наиболее насыщен бараний жир, который практически не содержит незаменимых кислот. Ценными пищевыми жирами являются рыбий жир и растительные масла, содержащие незаменимые жирные кислоты. Вопрос 34. Триацилглицеролы. Простые и смешанные. Физико-хим свойства жиров. Биолог. Роль Ацилглицеролы - сложные эфиры трёхатомного спирта глицерола и жирных кислот. Глицерол может быть связан с одной, двумя или тремя жирными кислотами, соответственно образуя моно-, ди- или триацилглицеролы (МАГ, ДАГ, ТАГ). Основную массу лигщдов в организме человека составляют триацилглицеролы - жиры. У человека с массой тела 70 кг в норме содержится до 10 кг жиров. Они запасаются в жировых клетках -- адипоцитах и используются при голодании как источники энергии. Моно- и диацилглицеролы образуются на промежуточных этапах распада и синтеза триацил-глицеролов. Атомы углерода в глицероле по-разному ориентированы в пространстве (рис. 8-2), поэтому ферменты различают их и специфически присоединяют жирные кислоты у первого, второго и третьего атомов углерода. Номенклатура и состав природных триацил-глицеролов. В молекуле природного жира содержатся разные жирные кислоты. Как правило, в позициях 1 и 3 находятся более насыщенные жирные кислоты, а во второй позиции - полиеновая кислота. В названии триацилглицерола перечисляются названия радикалов жирных кислот, начиная с первого углеродного атома глицерола, например пальмитоил-линоленоил-олеоилглицерол. Жиры, содержащие преимущественно насыщенные кислоты, являются твёрдыми (говяжий, бараний жиры), а содержащие большое количество ненасыщенных кислот - жидкими. Жидкие жиры или масла обычно имеют растительное происхождение . Из животных пищевых жиров наиболее насыщен бараний жир, который практически не содержит незаменимых кислот. Ценными пищевыми жирами являются рыбий жир и растительные масла, содержащие незаменимые жирные кислоты. . Различают простые и смешанные триацилглицерины. Простые — содержат остатки одинаковых ВЖК, а смешанные — остатки различных кислот. Все природные жиры не являются индивидуальными соединениями, а представляют собой смесь различных (как правило смешанных) три ацилглицеринов. ОБМЕН ТРИАЦИЛГЛИЦЕРОЛОВ Приём пищи человеком происходит иногда со значительными интервалами, поэтому в организме выработались механизмы депонирования источников энергии. Жиры - наиболее выгодная и основная форма депонирования энергии. Запасы гликогена в организме не превышают 300 г и обеспечивают организм энергией не более суток. Депонированный жир может обеспечивать организм энергией при голодании в течение длительного времени (до 7-8 нед). Синтез жиров активируется в абсорбтивный период и происходит в основном в жировой ткани и печени. Но если жировая ткань - место депонирования жира, то печень выполняет важную роль превращения части углеводов, поступающих с пищей, в жиры, которые затем секретируются в кровь в составе ЛПОНП и доставляются в другие ткани (в первую очередь, в жировую). Синтез жиров в печени и жировой ткани стимулируется инсулином. Мобилизация жира активируется в тех случаях, когда глюкозы недостаточно для обеспечения энергетических потребностей организма: в постабсорбтивный период, при голодании и физической работе под действием гормонов глюкагона, адреналина, соматотропина. Жирные кислоты поступают в кровь и используются тканями как источники энергии. Вопрос 35. Глицерофосфолипиды. Представители. Роль глицерофосфолипиды. Структурная основа глицерофосфолипидов - глицерол. Глицерофосфолипиды молекулы, в которых две жирные кислоты связаны сложноэфирной связью с глицеролом в первой и второй позициях; в третьей позиции находится остаток фосфорной кислоты, к которому, в свою очередь, могут быть присоединены различные заместители, чаще всего аминоспирты. Если в третьем положении имеется только фосфорная кислота, то глицерофосфолипид называется фосфатидной кислотой. Её остаток называют "фосфатидил"; он входит в название остальных глицерофосфолипидов, после которого указывают название заместителя атома водорода в фосфорной кислоте, например фосфатидилэтаноламин, фосфатидилхолин и т.д. Фосфатидная кислота в свободном состоянии в организме содержится в небольшом количестве), но является промежуточным продуктом на пути синтеза как три-ацилглицеролов, так и глицерофосфолипидов. У глицерофосфолипидов, как и у триацилгли-церолов, во второй позиции находятся преимущественно полиеновые кислоты; в молекуле фосфатидилхолина, входящего в структуру мембран, это чаще всего арахидоновая кислота. Жирные кислоты фосфолипидов мембран отличаются от других липидов человека преобладанием полиеновых кислот (до 80-85%), что обеспечивает жидкое состояние гидрофобного слоя, необходимое для функционирования белков, входящих в структуру мембран. Общая формула глицерофосфолипидов выглядит так: 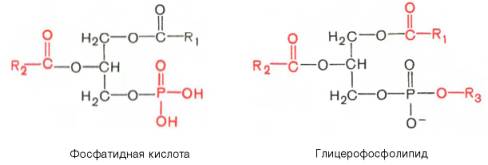 В отличие от триглицеридов в молекуле фосфатидилхолина одна из трех гидроксильных групп глицерина связана не с жирной, а с фосфорной кислотой. Кроме того, фосфорная кислота в свою очередь соединена эфирной связью с азотистым основанием –холином [НО—СН2—СН2—N+(CH3)3]. Таким образом, в молекуле фосфатидил-холина соединены глицерин, высшие жирные кислоты, фосфорная кислота и холин: 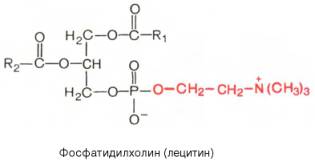 Фосфатидилэтаноламины. Основное различие между фосфатидилхоли-нами и фосфатидилэтаноламинами – наличие в составе последних азотистого основания этаноламина (HO—CH2—CH2—N+H3): 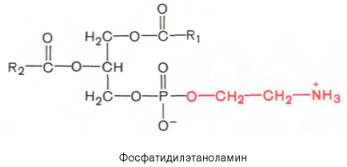 Из глицерофосфолипидов в организме животных и высших растений в наибольшем количестве встречаются фосфатидилхолины и фосфатидил-этаноламины. Эти 2 группы глицерофосфолипидов метаболически связаны друг с другом и являются главными липидными компонентами мембранклеток. Фосфатидилсерины. В молекуле фосфатидилсерина азотистым соединением служит остаток аминокислоты серина 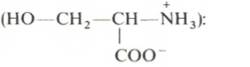 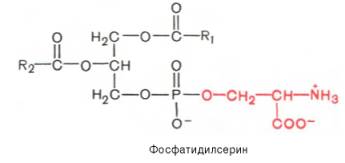 Фосфатидилсерины распространены гораздо менее широко, чем фос-фатидилхолины и фосфоэтаноламины, и их значение определяется в основном тем, что они участвуют в синтезе фосфатидилэтаноламинов. Фосфатидилинозитолы. Эти липиды относятся к группе производных фосфатидной кислоты, но не содержат азота. Радикалом (R3) в этом подклассе глицерофосфолипидов является шестиуглеродный циклический спирт инозитол: 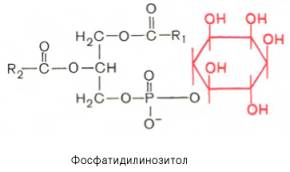 Фосфатидилинозитолы довольно широко распространены в природе. Они обнаружены у животных, растений и микроорганизмов. В животноморганизме найдены в мозге, печени и легких. | |||||||||||||||||||||||