|
|
ШПОРЫ БХ 2 ИТОГОВАЯ. Эндергонические процессы, протекающие с увеличением свободной энергии. Катаболические превращения
Вопрос 59. Биосинт х. в цитозоле клеток. 3 этапа
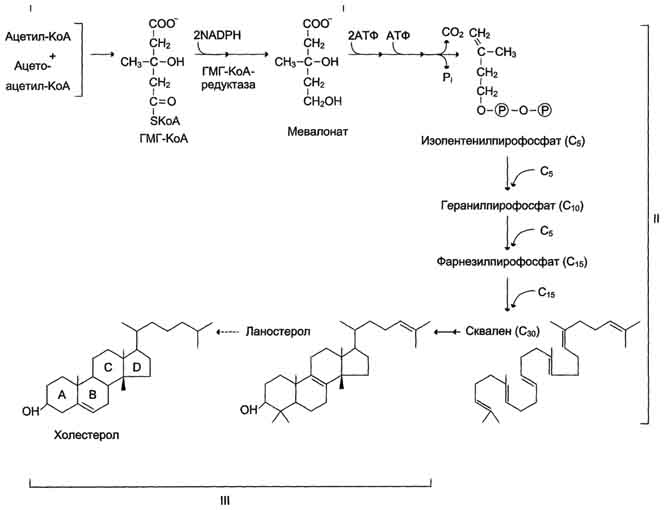
1. Образование мевалоната. Две молекулы ацетил-КоА конденсируются ферментом тиолазой с образованием ацетоацетил-КоА.. Фермент щдроксиметилглутарил-КоА-синтаза присоединяет третий ацетильный остаток с образованием ГМГ-КоА. Следующая реакция, катализируемая ГМГ-КоА-редуктазой, является регуляторной в метаболическом пути синтеза холестерола. восстановление ГМГ-КоА до мевалоната с использованием 2 молекул NADPH. Фермент ГМГ-КоА-редуктаза
2. Образование сквалена На втором этапе синтеза мевалонат превращается в пятиуглеродную изопреноидную структуру, содержащую изопентенилпирофосфат- геранилпирофосфат – фарнезилпирофосфата. Две молекулы фарнезилпирофосфата конденсируются с образованием сквалена
3. Образование холестерола сквален превращается в ланостерол. Далее происходит 20 последовательных реакций, превращающих ланостерол в холестерол. На последних этапах синтеза от ланостерола отделяется 3 атома углерода.
Регуляция синтеза холестерола. Регуляция активности ГМГ-КоА-редуктазы в печени. Холестерол и жёлчные кислоты снижают скорость транскрипции и, таким образом, синтез фермента. Инсулин стимулирует дефосфорилирование, а глюкагон - фосфорилирование ГМГ-КоА-редуктазы. Инсулин активирует 2 фосфатазы: киназы ГМГ-КоА-редуктазы* и фосфатазу, дефосфорилирующую непосредственно ГМГ-КоА-редуктазу. Глюкагон стимулирует фосфорилирование и инактивацию 2 фосфатаз и фосфорилирование и активацию киназы ГМГ-КоА-редуктазы.
Вопрос 60. Жк участвуют в переваривании жиров, эмульгируя их и делая доступными для действия панкреатической липазы. ЖК – производные холестерола с пятиуглеродной боковой цепью в положении 17, которая заканчивается карбоксильной группой. В организме человека синтезируются 2 жк: холевая (содержит 3 гидроксильные группы в положениях 3,7,12) и хендезоксихолиевая (содержит 2 гидроксильные группы в положениях 1 и 3). Жк не явл эффективными эмульгаторами в кишечнике, так как карбоксильные группы этих жк имеют рК
6. В печени эмульгирующие св-ва жк увеличиваются за счёт реакции коньюгации, в которой к карбоксильной группе присоединяется глицин или таурин, полностью ионизированные при рН кишечного сока.Эти производные называются солями жк и служат главными эмульгаторами жиров в кишечнике. Основная функция желчи – участие в переваривании и всасывании липидов. Эту функцию выполняют желчные кислоты, эмульгирующие жиры и таким образом обеспечивающие гидролиз жиров панкреатической липазой. Секреция желчи является также основным путем выведения избытка холестерина из организма. Он выводится как в виде желчных кислот, которые образуются в печени из холестерина, так и в свободном виде. Соотношение желчных кислот и холестерина должно быть приблизительно 12:1. при уменьшении образования желчи или увеличении количества в желчи холестерина последний выпадает в осадок. Этот густой маслянистый осадок пропитывается солями кальция, желчными пигментами и со временем превращается в камни.
Избыточная секреция холестерина с желчью – один из механизмов камнеобразования в желчном пузыре.
Пониженная секреция желчных кислот и фосфолипидов – еще одна причина камнеобразования.
1) уменьшение резерва желчных кислот;
2) усиленное превращение холевой кислоты в дезоксихолевую.
Первое может быть следствием ускоренного прохождения первичных желчных кислот из тонкой кишки в толстую, второе – усиленного дегидроксилирования холевой кислоты и ускоренного всасывания образующейся при этом дезоксихолевой кислоты.
В литогенной желчи ускорено образования мелких кристалликов моногидрата холестерина. Именно этой особенностью литогенная желчь отличается от нормальной. К предрасполагающим этому факторам можно отнести гликозаминогликаны. Препятствующим факторам образованию зародышей кристаллизации относят апопротеины А1 и А 2 и некоторые гликопротеиды.
Вопрос 61. Х. транспортируется кровью только в составе ЛПНП и ЛПВП. ЛП обеспечивают поступление в ткани экзогенного х., определяют потоки х. между органами и выведение избытка х. из организма.
ЛПНП - основная транспортная форма х., в которой он доставляется в ткани. Около 70% х. и его эфиров в крови находится в составе ЛПНП. Из крови ЛПНП поступают в печень (до 75%) и другие ткани, которые имеют на своей поверхности рецепторы ЛПНП.
ЛПВП вып. 2 осн.ф-и: поставляют апопротеины другим ЛП в крови и участвуют в "обратном транспорте х.". ЛПВП синтезируются в печени и в небольшом количестве в тонком кишечнике. В печени в ЛПВП включаются апопротеины А, Е, С-II, фермент ЛХАТ. В крови апоС-II и апоЕ переносятся с ЛПВП на ХМ и ЛПОНП. Для переноса х. На поверхности ЛПВП находится фермент ЛХАТ, кот превращает х., имеющий гидроксильную группу, выступающую на поверхность липопротеинов или мембран клеток, в эфиры холестерола. Гидрофобная молекула, эфира холестерола перемещается внутрь ЛПВП. образом, часть эфиров холестерола переносится на ЛПОНП, ЛППП, а ЛПВП3 за счёт накопления триацилглицеролов увеличиваются и превращаются в ЛПВП2. ЛПОНП под действием ЛП-липазы превращаются сначала в ЛППП, а затем в ЛПНП. ЛПНП и ЛППП захватываются клетками через рецепторы ЛПНП. Липопротеины различаются и по участию в атерогенезе. Атерогенность липопротеинов частично зависит от размера частиц. Самые мелкие липопротеины, такие как ЛПВП, легко проникают в стенку сосуда, но также легко ее покидают, не вызывая атеросклероз. ЛПНП – наиболее атерогенные липопротеины крови. достаточно малы, чтобы проникать в стенку сосуда, и легко задерживаются в сосудистой стенке.
Рецептор ЛПНП - сложный белок, состоящий из 5 доменов и содержащий углеводную часть Рецепторы ЛПНП синтезируются в ЭР и аппарате Гольджи, а затем экспонируются на поверхности клетки, в специальных окаймлённых ямках. Выступающий на поверхность N-концевой домен рецептора взаимодействует с белками апоВ-100 и апоЕ; В регуляции синтеза рецепторов ЛПНП участвуют гормоны: инсулин и трийодтиронин, половые гормоны. Они увеличивают образование рецепторов ЛПНП, а глюкокортикоиды уменьшают
Вопрос 62. Концентрация холестерола в крови взрослых людей составляет 200±50 мг/дл (5,2±1,2 ммоль/л) и, как правило, увеличивается с возрастом. Превышение нормальной концентрации холестерола в крови называют гиперхолестеролемией.
Дислипопротеинемии: наследственная недостаточность ЛП-липазы, 2- семейная комбинированная гиперлипидемия, нарушение удаления остаточных липопротеинов из крови, 3- семейная гипертриглицеролемия.Гиперхолестеролемия развивается вследствие избыточного поступления холестерола с пищей.Правильное питание в течение всей жизни - важнейший фактор профилактики гаперхолестеролемии. Доказана корреляция между увеличением концентрации холестерола в плазме крови и смертностью от заболеваний ССС - инфаркта миокарда и инсульта, развивающихся в результате атеросклероза (рис. 8-74).
Ген рецептора ЛПНП: структура и типы мутаций
Ген рецептора ЛПНП находится в хромосоме 19 и состоит из 18 экзонов Различные группы экзонов кодируют различные домены в составе этого белка. Мутации в этом гене подробно изучены и разделены на 4 класса.
Первый класс приводит к полному отсутствию рецептора; второй класс мутаций - рецептор синтезируется, но не может транспортироваться на поверхность клетки; третий класс мутаций - рецептор транспортируется на поверхность клеток, но не связывает ЛПНП; четвёртый класс мутаций - рецептор связывает ЛПНП, но не происходит эндоцитоз. Изменения структуры рецепторов ЛПНП в результате всех типов мутаций приводит к гиперхолестеролемии; так как ЛПНП не захватываются клетками, и холестерол в составе ЛПНП накапливается в крови. Важным лечебным и профилактич фактором, снижающим риск развития гиперхолестеролемии и атеросклероза, является гипокалорийная и гипохолестериновая диета. Поступление холестерола с пищей не должно превышать 300 мг/сут. Растительная пища не содержит холестерола, должна составлять основу рациона. К лечебным и профилактическим факторам относят обогащение пищи полиеновыми жирными кислотами семейства ω-3, уменьшающими риск тромбообразования. Антиоксиданты и Витамины С, Е, А поддерживают нормальную структуру липидов ЛПНП и их метаболизм. Лечение гиперхолестеролемии, как правило, комплексное. Один из принципов лечения - "размыкание" цикла энтерогепатической циркуляции жёлчных кислот. Используют лекарства типа холестирамина - полимера, который в кишечнике адсорбирует жёлчные кислоты, выделяется с фекалиями и уменьшает возврат жёлчных кислот в печень. Наиболее эффективные препараты, применяемые при лечении атеросклероза, - ингибиторы ГМГ-КоА-редуктазы. Эти препараты - антибиотики, например мевакор, в печени эффективно ингибируют регуляторный фермент биосинтеза холестерола. фибраты - ускоряют катаболизм ЛПОНП, активируя ЛП-липазу.
Вопрос 63. Эйкозаноиды - биологически активные вещества, синтезируемые большинством клеток из полиеновых жирных кислот, содержащих 20 углеродных атомов. Эйкозаноиды, включающие в себя простагландины, тромбоксаны, лейкотриены и ряд других веществ, - высокоактивные регуляторы клеточных функций. регулируют тонус ГМК и вследствие этого влияют на АД, состояние бронхов, кишечника, матки, регулируют секрецию воды и натрия почками, влияют на образование тромбов,участвуют в развитии воспалительного процесса, происходящего после повреждения тканей или инфекции. Главный субстрат для синтеза эйкозаноидов у человека - арахидоновая кислота.Простагландины обозначают символами, например PG А, где PG обозначает слово "простагландин", а буква А обозначает заместитель в пятичленном кольце в молекуле эйкозаноида. Каждая из указанных групп простагландинов состоит из 3 типов молекул, отличающихся по числу двойных связей в боковых цепях. Число двойных связей обозначают нижним цифровым индексом, например, PG Е2.
Тромбоксаны. В отличие от простагландинов, тромбоксаны синтезируются только в тромбоцитах и стимулируют их агрегацию при образовании тромба.
синтез простагландинов и тромбоксанов
Синтез простагландинов. Фермент, катализирующий первый этап синтеза простагландинов, называется PG Н2 синтазой и имеет 2 каталитических центра. Один из них называют циклооксигеназой, другой - пероксидазой. В активном центре циклооксигеназы находится тирозин, в активном центре пероксидазы - гем. В организме имеются 2 типа циклооксигеназ (PG Н2 синтаз). Циклооксигеназа 1 - конститутивный фермент, синтезирующийся с постоянной скоростью. Синтез циклооксигеназы 2 увеличивается при воспалении и индуцируется соответствующими медиаторами - цитокинами. Оба типа циклооксигеназ катализируют включение 4 атомов кислорода в арахидоновую кислоту и формирование пятичленного кольца. В результате образуется нестабильное гидропероксидпроизводное PG G2. Гидропероксид у 15-го атома углерода быстро восстанавливается до гидроксильной группы пероксидазой с образованием PG Н2.
Лейкотриены также образуются из эйкозаноевых кислот они имеют 3 сопряжённые двойные связи, хотя общее число двойных связей в молекуле больше. Липоксигеназный путь синтеза, начинается с присоединения молекулы кислорода к одному из атомов углерода у двойной связи, с образованиемГПЭ-ТЕ. Далее гидропероксиды превращаются в ГЭТЕ).
Структура и синтез лейкотриенов и ГЭТЕ
Синтез лейкотриенов идёт по пути, отличному от пути синтеза простагландинов, и начинается с образования гидроксипероксидов - гидропероксидэйкозатетраеноатов (ГПЭТЕ). Эти вещества или восстанавливаются с образованием гидроксиэйкозатетроеноатов (ГЭТЕ) или превращаются в лейкотриены или липоксины.
Вопрос 64. Липиды мембран. Основные липидные компоненты биологические мембраны - фосфолипиды, гликолипиды и стерины.
В клетках млекопитающих плазматические мембраны обогащены х. и гликосфииголипидами, Наиболее распространенные липидыв большинстве мембран клеток млекопитающих фосфатидилхолин и сфингомиелин. Дифосфатидилглицерин в значительных количествах присутствует только в мембранах митохондрий. В плазматических мембранах содержание фосфатидилсерина обычно больше, чем фосфатидилинозита. В мембранах миелина широко представлены цереброзиды. стерины мембран растительных клеток - ситостерин и стигмастерин, мембран грибов и дрожжей - эргостерин и зимостерин.
Липиды - основной строительный материал, из которого формируются клеточные мембраны. они участвуют также в регуляции важнейших мембранных процессов.
Нейтральные липиды, составляющие до 30% (по массе) от всех липидов, включают глицериды, холестерин и эфиры холестерина. Главной фракцией является холестерин.
Гликолипиды составляют около 10% мембранных липидов и являются гликосфинголипидами. Они содержат ненасыщенный аминоспирт сфингозин, длинноцепочечную ненасыщенную жирную кислоту и полярные углеводные группы. играют роль в клеточной адгезии и узнавании различных молекул, регуляции электрической проводимости, защитной функции и т.д.
Расщепление фосфолипидов идет гидролитическим путем при участии ферментов фосфолипаз, поступающих в двенадцатиперстную кишку с соком поджелудочной железы. Фосфолипаза А1 катализирует расщепление сложноэфирной связи между ацилом и первым атомом углерода глицерола.Фосфолипаза А2 катализирует гидролиз сложноэфирной связи между ацилом и вторым атомом углерода глицерола. Фосфолипаза С катализирует гидролитический разрыв связи между третьим атомом углерода глицерола и остатком фосфорной кислоты, а фосфолипаза Д ДД сложноэфирной связи между остатком фосфорной кислоты и остатком аминоспирта.
В результате действия этих четырех ферментов фосфолипиды расщепляются до свободных жирных кислот, глицерола, фосфорной кислоты и аминоспирта.
Вопрос65. Влияние инсулина на метаболизм глюкозы. Около 50% глюкозы используется в процессе Гликолиза, 30-40% превращается в жиры и около 10% накапливается в форме гликогена. Общий результат стимуляции этих процессов - снижение концентрации глюкозы в крови.
Влияние инсулина на метаболизм жиров. В печени и жировой ткани инсулин стимулирует синтез жиров, обеспечивая получение для этого процесса необходимых субстратов (ацетил-КоА, α-глицерофосфат и NADPH) из глюкозы. В адипоцитах инсулин активирует ацетил КоА-карбок-силазу и ЛП-липазу и индуцирует синтез синта-зы жирных кислот, ацетил-КоА-карбоксилазы и ЛП-липазы. Инсулин в жировой ткани тормозит мобилизацию жиров. снижается концентрация жирных кислот, циркулирующих в крови. Основную роль в поддержании энергетического гомеостаза играют гормоны инсулин и глюкагон. а также другие контринсулярные гормоны - адреналин, кортизол, йодтиронины и соматотропин. Инсулин и глюкагон играют главную роль в регуляции метаболизма при смене абсорбтивного и постабсорбтивного периодов и при голодании.
Вопрос 66. Сахарный диабет — хронический метаболический синдром, характеризующийся гипергликемией, глюкозурией и связанными с ними нарушениями обмена веществ. Развивается вследствие абсолютной или относительной (нарушение взаимодействия с клетками-мишенями) недостаточности гормона инсулина и приводит к нарушению углеводного, жирового и белкового обмена.
I. Сахарный диабет 1-го типа
А. Аутоиммунный
Б. Идиопатический
II. Сахарный диабет 2-го типа
III. Другие специфические типы сахарного диабета
Тест толерантности к глюкозе – определение содержания глюкозы в крови. Кровь из пальца берут до приема глюкозы и ч/з 1 и 2 часа после приема 75 г глюкозы. У здорового человека натощак показатель содержания глюкозы <5,6, ч\з 2 ч после приема глюкозы <7,7. При СД натощак >6,7, >11,1. На основе полученных результатов составляют гликемические кривые.
Клиническое значение: При СД, панкреатите, воспалениях, инфекциях, гиперфункции щитовидной железы – высокий подьем глик.кривой и медленное уменьшение до прежнего уровня. Глюкозы при подобных заболев. переходит в мочу. |
|
|
 Скачать 2.83 Mb.
Скачать 2.83 Mb.