УЧЕБНИКгенетика. Генетика изучает процессы преемственности жизни на молекулярном, клеточном, организменном и популяционном уровнях
 Скачать 6.93 Mb. Скачать 6.93 Mb.
|

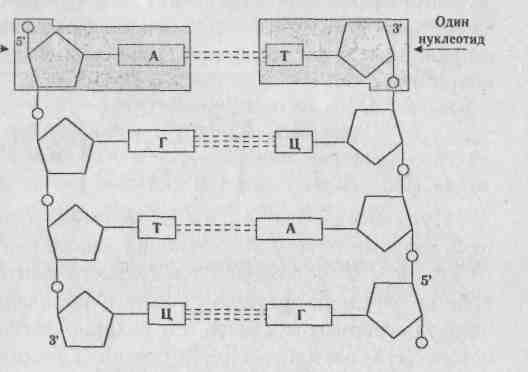
МОЛЕКУЛЯРНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ 2.1. Генетическая роль ДНК и РНК 2.1.1. ДНК — носитель наследственной информации «Значение ДНК столь велико, что никакое знание о ней не будет полным». Ф.Крик. ДНК — дезоксирибонуклеиновая кислота — биологическая макромолекула, носитель генетической информации во всех эукариотических и прокариотических клетках и во многих вирусах. В 1928 г. Ф.Гриффит обнаружил у пневмококков явление трансформации (преобразование свойств бактерий). Он показал, что клетки невирулентных штаммов бактерий (шероховатые без капсул) приобретают свойства вирулентных (гладких с капсулами) штаммов, убитых нагреванием. Природа трансформирующего агента была установлена Эвери, Мак-Леодом и Мак-Карти в 1944 г., им оказалась ДНК. Так открытие и изучение трансформации доказало роль ДНК как материального носителя наследственной информации, (рис. 2.1). 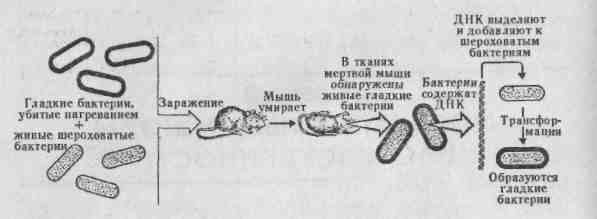 Рис. 2.1. Трансформирующий фактор — это ДНК- Трехмерная модель пространственного строения двухцепочечной ДНК была описана в апрельском журнале Nature в 1953 г. Дж. Уотсоном, Френсисом Криком и Морисом Уилкинсом. Эти исследования легли в основу молекулярной биологии, изучающей основные свойства и проявления жизни на молекулярном уровне. Структура ДНК — полимер, структурной единицей которого является нуклеотид (рис. 2.2). Нуклеотид состоит из азотистого основания пу-ринового: аденин (А) или гуанин (Г) или пирими-динового: цитозин (Ц) или тимин (Т), углевода дезоксирибозы (пятиуглеродное сахарное кольцо) и остатка фосфорной кислоты (НРО |

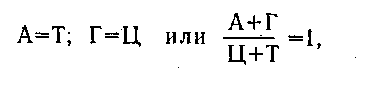
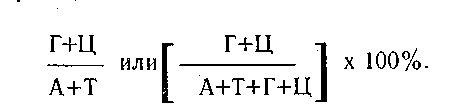
| Тип спирали | Число пар основа- нии на виток | Угол враще- ния одной пары, градусы | Рассто- яние между парами в А° 10"9 | Диаметр спирали А° 10"9 | Напра- вление спирали |
| А | 11 | +32.7 | 2.56 | 23 | правая |
| В | 10 | +36.0 | 3.38 | 19 | правая |
| С | 9.3 | +38.6 | 3.32 | 19 | правая |
| Z | 12 | -30.0 | 3.71 ' | 18 | правая и левая |
Знание структуры и функции ДНК необходимо для понимания сути некоторых генетических процессов, которые являются матричными. Было ясно, что сама ДНК не может играть роль матрицы при синтезе белков из аминокислот, т.к. почти вся она находится в хромосомах, расположенных в ядре, в то время как большинство, если не все, клеточные белки синтезируются в цитоплазме. Таким образом, генетическая информация, заклю- -ченная в ДНК, должна передаваться какой-то промежуточной молекуле, которая транспортировалась бы в цитоплазму и участвовала в синтезе полипептидных цепей. Предположение о том, что такой промежуточной молекулой может быть РНК, стало всерьез рассматриваться сразу, как только была открыта структура двойной спирали ДНК. Во-первых, клетки, синтезирующие большое количество белка, содержали много РНК. Во-вторых, еще более важным казалось то, что сахарофос-фатные «скелеты» ДНК и РНК чрезвычайно сходны и было бы легко представить себе, как происходит синтез одиночных цепей РНК на одноцепочеч-ной ДНК с образованием нестабильных гибридных молекул, одна цепь которых представлена ДНК, а другая РНК. Взаимоотношения ДНК, РНК и белка в 1953 г. были представлены в виде следующей схемы:
репликация ДНК..... транскрипция - -----> РНК... трансляция......-> белок,
где одиночные цепи ДНК служат матрицами при синтезе комплементарных молекул ДНК (репликация). В свою очередь, молекулы РНК служат матрицами для последовательного соединения аминокислот с образованием полипептидных цепей белков в процессе трансляции, названном так потому, что «текст», написанный на «языке» нуклеотидов, переводится (транслируется) на «язык» аминокислот. Группа нуклеотидов, кодирующая одну аминокислоту, называется кодоном.
2.1.2. Строение и функции РНК
РНК — рибонуклеиновая кислота, имеет много общего со структурой ДНК, но отличается от нее рядом признаков:
углеводом РНК, к которому присоединяются пуриновые или пиримидиновые основания и фосфатные группы, является рибоза;
в состав РНК, как и в состав ДНК, входят азотистые основания аденин, гуанин и цитозин. Но РНК не содержит тимина, его место в молекуле РНК занимает урацил;
РНК — одноцепочечная молекула;
так как молекула РНК одноцепочечная, то правило Чаргаффа, установленное для ДНК, может не выполняться по равенству содержания оснований.
Рибонуклеиновые кислоты (РНК), присутствующие в клетках как про- так и эукариот, бывают трех основных видов: матричные РНК (мРНК), рибосомные РНК (рРНК) и транспортные РНК (тРНК).
Матричные РНК (информационная РНК, мессенджер-РНК) выполняют функцию матриц белкового синтеза (см. «Транскрипция»). В ядре клеток эукариот содержится РНК четвертого типа гетерогенная ядерная РНК(гяРНК), которая является точной копией (транскриптом) соответствующей ДНК. Процесс транскрипции осуществляется в ядре на ДНК, гяРНК после созревания будет служить матрицей для синтеза белка в цитоплазме.
Молекулы тРНК узнают в цитоплазме соответствующий триплет (кодон в мРНК) и переносят нужную аминокислоту к растущей полипептидной цепи. Узнавание кодона в мРНК осуществляется с помощью трех последовательных оснований в тРНК, называемых антикодонами. Аминокислотный остаток может присоединятся к 3' — концу молекулы тРНК. Считают, что для каждой аминокислоты имеется, по крайней мере, одна тРНК. Структура тРНК представлена на рис. 2.3. Молекула тРНК содержит около 75 нуклеотидов, ковалентно связанных друг с другом в линейную цепочку. Иногда эту структуру называют «клеверным листом», конфигурация которого возникает благодаря нескольким внутрицепочечным комплементарным участкам. Молекулы всех видов тРНК имеют 4 основных плеча. Акцепторное плечо заканчивается последовательностью ЦЦА (5—3'). Через 3' происходит связывание с карбоксильной группой аминокислоты. Остальные плечи тоже состоят из стеблей, образованных комплементарными парами оснований и петель из неспарен-ных оснований. Антикодоновое плечо узнает нук-леотидный триплет (кодон).
Рибосома состоит из большой и малой субъединиц. Для эукариот рибосома состоит из большой субъединицы с молекулярной массой 2,8 х106 (60S) и малой, имеющей молекулярную массу 1,4x10"6 (40S), где S (Сведберг)— единица измерения коэффициента седиментации (мера массы макромолекулы). Эти субчастицы могут диссоциировать на белок и рРНК. Весовое соотношение рРНК: белок для эукариот составляет 1:1. Все субчастицы рибосом состоят из рРНК, которые синтезируют-ся в структуре ядрышек. В цитоплазме рибосомы упаковываются с рибосомными белками, приобретая достаточную устойчивость, и способны осуществлять большое число циклов трансляции.
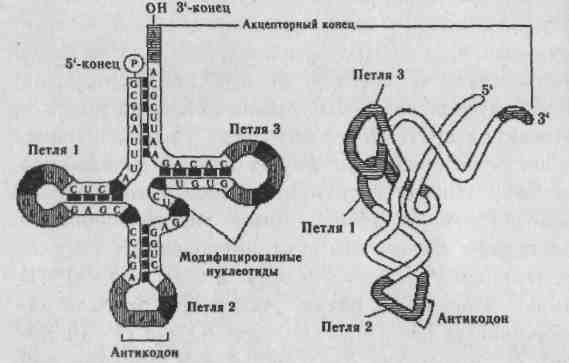
Рис. 2.3. Строение тРНК.
В 1970-х годах Сидней Альтман и Томас Чек сделали открытие, противоречащее традиционным представлениям. Они показали, что РНК не только является переносчиком информации с ДНК, но и выступает в роли катализатора в ряде клеточных процессов. Каталитическая способность РНК позволила использовать эту молекулу для различных биотехнологических целей, в частности для борьбы с вирусными инфекциями.
2.2. Генетический код
До расшифровки генетического кода невозможно было понять, каким образом кодируется наследственная информация. Расшифровка генетического кода позволила понять механизм синтеза белка и связать между собой дефекты определенных белков человека и наследственные заболевания, а также создала необходимые предпосылки для диагностики и лечения таких заболеваний.
Генетический код — единая система записи наследственной информации в молекулах нуклеиновых кислот в виде последовательности нуклеоти-дов. Генетический код основан на использовании алфавита, состоящего всего из четырех букв-нуклео-тидов, отличающихся азотистыми основаниями: А, Т, Ц, Г. Попытки расшифровки генетического кода были предприняты в 1954 г. Г.Гамовым. Основные свойства кода триплетность и вырожденность выявили в 1961 г. Ф.Крик и С.Бреннер.
Поскольку в белках встречается 20. различных аминокислот, то каждая не может кодироваться одним нуклеотидом (в ДНК всего 4 типа нуклеоти-дов), т.к. 16 останутся незакодированными. Не могут «слова» генетического кода состоять и из 2 «букв», т.к. будут закодированы всего 16 аминокислот. Значит, наименьшая возможная длина «слова», определяющая аминокислоту, — 3 нуклеотида. Число • возможных триплетов нуклеотидов равно 64.
В 1961 г. была впервые дешифрована первая триплетная последовательность. Система, содержащая искусственную мРНК, состоящую только из урациловых нуклеотидов, синтезировала полипептидную цепь, состоящую только из фенилаланина (в ДНК кодом для нее должен быть комплементарный триплет нуклеотидов — AAA). Это сделали двое ученых М.Ниренберг и Г.Маттеи. К 1965 г. был расшифрован полностью весь генетический код (рис. 2.4). Из 64 кодонов три кодона УАГ, УАА, УГА не кодируют аминокислот, они были названы нонсенс-кодонами. Позднее было показано, что они являются терминирующими кодонами.
В настоящее время определение нуклеотидных последовательностей ДНК и РНК проводится с помощью специального метода — секвенирования.
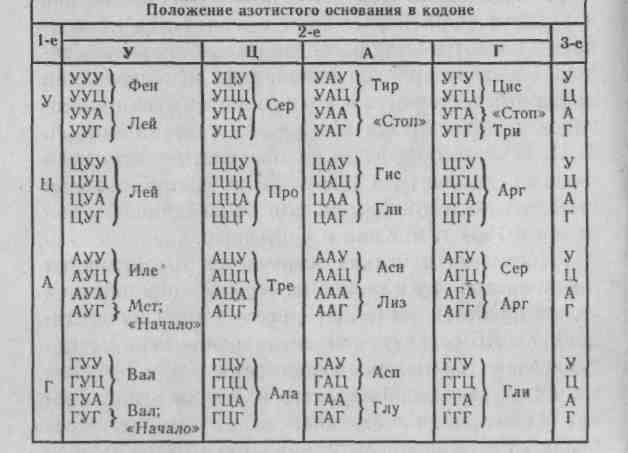
Рис. 2.4. Генетический код.
Свойства генетического кода
1. Генетический код триплетен. Триплет (кодон) — последовательность трех нуклеотидов, кодирующая одну аминокислоту.
Шутливая ода-четверостишие, посвященная генетическому коду, написана генетиками. Каждое слово состоит из трех букв, как триплет, кодирующий одну аминокислоту, из трех нуклеотидов:
Наш код так мал,
Так лих наш код.
Был дым, был — пал,
Нет, жив наш род!
2. Вырожденность генетического кода обусловлена тем, что одна аминокислота может кодироваться несколькими триплетами (вспомним, что аминокислот 20, а триплетов — 64), исключение составляют метионин и триптофан, которые кодируются только одним триплетом. Три триплета УАА, УАГ, УГА — это стоп-сигналы (терминирующие кодоны), прекращающие синтез полипептидной цепи. Триплет, соответствующий метионину (АУГ), выполняет функцию инициирования (возбуждения) считывания и не кодирует аминокислоту, если стоит в начале цепи ДНК..
3. Однозначность — каждому данному кодону соответствует одна и только одна определенная аминокислота. Следует отчетливо понимать принципиальное отличие двух важнейших свойств — вырожденности и однозначности, одновременно присущих генетическому коду.
4. Генетический код не перекрываем — процесс считывания генетического кода не допускает возможности перекрывания кодонов. Начавшись на определенном кодоне, считывание следующих идет без пропусков вплоть до нонсенскодонов. Таким образом, генетический код не содержит знаков пунктуации.
5. Генетический код универсален, т.е. вся информация в ядерных генах для всех организмов, обладающих разным уровнем организации (например, бабочка, ромашка, рак, лягушка, удав, орел, человек), кодируется одинаково.
При считывании информации положение первого основания кодона (триплета) определяет рамка считывания. Число возможных рамок считывания равно трем, поскольку генетический код триплетен (рис. 2.5). Обычно функциональный белок синтезируется по одной рамке считывания.
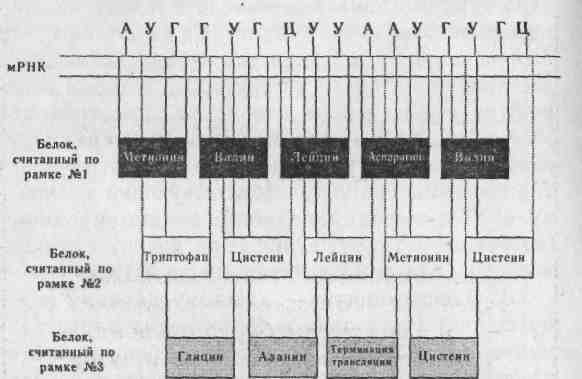
Рис. 2.5. Изменения в белке, происходящие при сдвиге рамки считывания. Нуклеотидную последовательность можно прочесть по одной из трех рамок считывания, причем каждый раз будет получаться новый белок.
Иногда могут происходить изменения рамки считывания, связанные с выпадением или добавлением одного или нескольких нуклеотидов. При последующей сборке белка в нем будет нарушена последовательность аминокислот. Это называется мутацией со сдвигом рамки.