УЧЕБНИКгенетика. Генетика изучает процессы преемственности жизни на молекулярном, клеточном, организменном и популяционном уровнях
 Скачать 6.93 Mb. Скачать 6.93 Mb.
|

|
Глава 3 МАТРИЧНЫЕ ПРОЦЕССЫ В КЛЕТКЕ Существуют три типа матричных процессов в клетках: репликация, транскрипция и трансляция. Кратко остановимся на них. 3.1. Репликация ДНК Основное функциональное значение процесса репликации ДНК заключается в снабжении потомства генетической информацией, которая должна передаваться полностью и с очень высокой точностью. Репликация — удвоение ДНК, происходящее в синтетическую (S) стадию интёрфазы перед каждым делением клетки. Информация, необходимая для репликации ДНК, заложена в ее структуре. Поскольку нити ДНК комплементарны друг другу, т.е. основания в парах дополняют друг друга, каждая цепь автоматически поставляет информацию для образования недостающей цепи. В 1957 г. Дельбрук и Стент сформулировали три альтернативные гипотезы репликации ДНК в клетках эукариот: Консервативная репликация. Исходная двухцепочечная молекула ДНК служит матрицей для 2. Основы генетики человека образования совершенно новой двухцепочечной молекулы, нацело достраивающейся на исходной. При этом одна из дочерних клеток получит исходную ДНК, а другая вновь синтезированную ДНК. Полуконсервативная репликация. Две нити ДНК расплетаются (как застежка-молния). Каждая цепь служит матрицей для образования новой. При репликации молекула ДНК постепенно разделяется специальным ферментом на две половины в продольном направлении. По мере того, как открываются нуклеотиды разделяемой молекулы, к ним тут же присоединяются свободные нуклеотиды, ранее синтезированные в цитоплазме. В результате каждая половинная спираль снова становится целой, и вместо одной молекулы получаются две, в результате чего хромосома становится двухроматидной. Дисперсионная репликация. Исходная ДНК распадается на короткие разной длины фрагменты, используемые в качестве матриц для построения фрагментов двух новых двойных спиралей, которые затем воссоздаются в единую структуру молекулы. Образованные молекулы ДНК содержат старые и новые фрагменты. Позже М.Мезельсон и Ф.Сталь, используя авторадиографический метод, показали, что полуконсервативный метод репликации характерен для всех эукариот и большинства прокариот. Лишь только некоторые формы вирусов способны к дисперсионной и консервативной формам репликации. Процесс репликации ДНК весьма сложен, но протекает аналогично у про- и эукариот, отличаясь участвующими ферментами, скоростью и направлением репликации, количеством точек репликации (числом репликативных вилок-репликонов). Скорость репликации у эукариот осуществляется довольно медленно — 50 пар оснований в 1 с, а у E.coli 1700 пар/с, но малая скорость репликации компенсируется множеством репликонов и двунаправленностью репликации. В каждой репликативной вилке ДНК расплетена и находится в одноцепочечной форме. Репликативная вилка перемещается вдоль молекулы, расплетая ДНК, пока не дойдет до точки окончания синтеза (рис. 3.1). 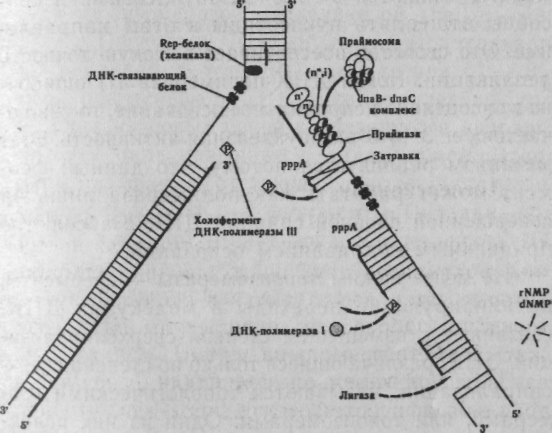 рис. 3.1. Строение репликативной вилки E.Coli(А.Корнберг, 1984). В 1955 г. А.Корнберг и его коллеги из Стенфордского университета открыли фермент, который обеспечивает репликацию ДНК, и назвали его полимеразой. Полимераза присоединяет компле-ментарный нуклеотид к матричной цепи. Например, в матричной цепи находится нуклеотид А — полимераза присоединяет к нему нуклеотид Т, если в матричной цепи встречается Г-нуклеотид, полимераза к нему присоединяет Ц-нуклеотид. На современном этапе среди ферментов, участвующих в синтезе ДНК, выделены ДНК-полимеразы I, II, III, обладающие 5'-> 3'полимеразной активностью. Кроме того, все три фермента проявляют способность деградировать ДНК. Полимеразы являются 3'—> 5' экзонуклеазами и способны отщеплять нуклеотиды в этом направлении. Это свойство обеспечивает высокую точность репликации. Когда ДНК-полимераза III ошибочно присоединит неправильное основание, то «включается» ее 3'—> 5'экзонуклеазная активность. Этот механизм реализуется потому, что данный фермент может работать как полимераза лишь на ' совершенной двойной спирали ДНК с абсолютно правильным спариванием оснований. Не менее важны топоизомеразы — ферменты, катализирующие переходы в молекулах ДНК, связанные с изменением степени сверхспирализа-ции. ДНК, различающиеся только по степени сверх-спирализации, называются топологическими изомерами, или топоизомерами. Одни из них релак-сируют сверхспирали, а другие — приводят к образованию, в них сверхвитков. В 1967 г. была открыта ДНК-гираза, которая переводит двухцепочечную ДНК в состояние отрицательной сверхспи-рализации. Это необходимо для снятия положительных сверхвитков, возникающих при репликации из-за раскручивания двойной спирали. Сверхспирали регулируют активность ДНК, а4 степень сверхспирализации контролируется ферментами то-поизомеразами-П (для эукариот)или гиразами. Инициация репликации начинается с образования сверхспиралей у ДНК. Этот процесс реализуется в присутствии ДНК-гиразы и АТФ. Если вывести из стоя гиразу, репликация прекращается. Сверхспирализация ДНК происходит лишь в тех случаях, когда не нарушена целостность структуры ДНК. Сверхспираль сразу же раскручивается топоизомеразой — хеликазой (helix — спираль). Поскольку ДНК-полимеразы катализируют репликацию только в направлении 5' -> 3', а цепи родительской ДНК антипараллельны, только одна из новых цепей синтезируется непрерывно. Эта цепь называется лидирующей. Вторая цепь, называемая отстающей, синтезируется в виде фрагментов ДНК — фрагменты Оказаки, которые у эукариот имеют последовательность 100— 200 нуклеотидов. Эти фрагменты лигируются (сшиваются) полинуклеотидлигазами, и образуется непрерывная вторая цепь. Этот процесс называется созреванием. Синтез каждого фрагмента Оказаки (3' —> 5') начинается на маленьком фрагменте РНК (около 10—60 нуклеотидов), который удаляется еще до окончания считывания фрагмента. Это так называемая затравка, или праймер. При созревании РНК затравка удаляется, как с 5' — конца лидирующей цепи, так и 5'— концов фрагментов Оказаки, с помощью ДНК-полимеразы I, действующей как 3' -» 5' экзонуклеаза, ДНК-лигаза соединяет в нужном порядке фрагменты ДНК (рис. 3.2). 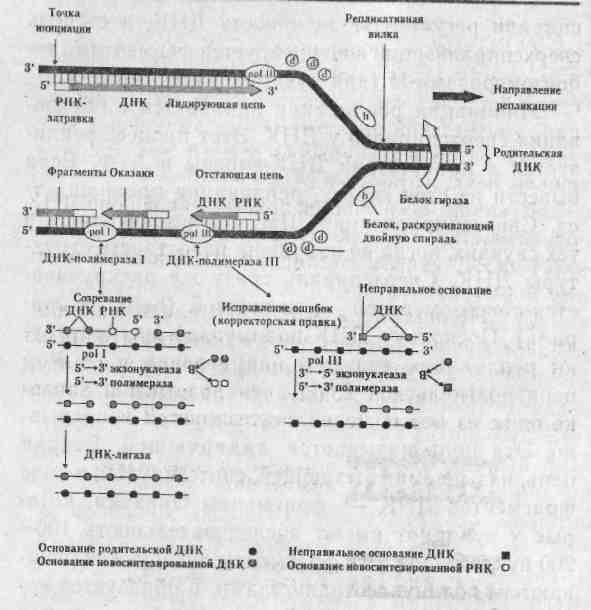 Рис. 3.2. Репликация у E.Coli (Э.Рис, М.Стернберг, 1988). 3.2. Репарация ДНК В любой клетке человека под влиянием различных факторов в ДНК ежедневно происходят тысячи случайных изменений, а за год в каждой клетке накапливается лишь очень небольшое число стабильных изменений нуклеотидной последовательности ДНК. Среди множественных случайных замен оснований в ДНК лишь одна на тысячу приводит к возникновению мутации. Все остальные повреждения очень эффективно ликвидируются в процессе репарации ДНК- Механизм репарации («залечивание» повреждений ДНК) основан на том, что молекула ДНК имеет две копии генетической информации — по одной в каждой из нитей молекулы. Основной путь репарации включает три этапа:
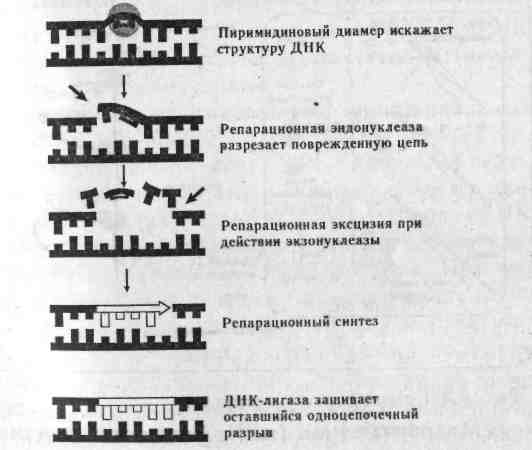 Рис. 3.3. Этапы вырезания и репарации поврежденного участка ДНК (Айала, 1988). Нарушение репарации у людей, пораженных пигментной ксеродермой (повышенная чувствительность к солнечному свету), приводит к раку кожи. Активность УФ-эндонуклеазы, необходимая для репарации, при этом отсутствует. Если подавляются репарационные системы, то мутагенез усиливается. 3.3. Биосинтез белка Транскрипция (переписывание) — синтез на ДНК-матрице мРНК (первичного продукта гена), осуществляющийся в ядре на смысловой нити ДНК, находящейся в релаксированном (деспирализованном) состоянии (рис. 3.4). Это первый этап белкового синтеза. . 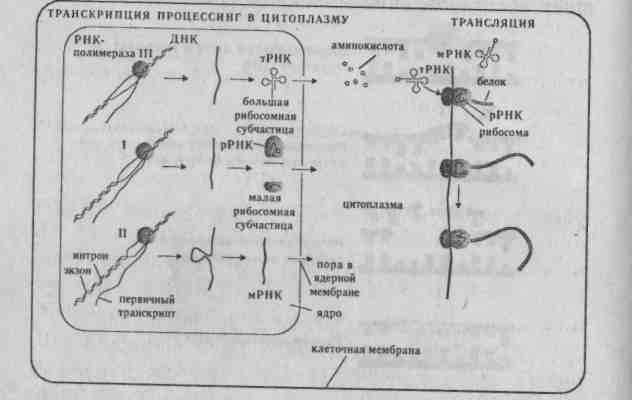 Рис. 3.4. Синтез белка у эукариот. Матричная РНК (мРНК) содержит генетическую инструкцию по синтезу определенного полипептида и переносит ее к белоксинтезирующему аппарату клетки, находящемуся в рибосомах цитоплазмы клеток. Синтез мРНК имеет стадии: инициации, элонгации и терминации аппарату клетки, находящемуся в рибосомах цитоплазмы клеток. Синтез мРНК имеет стадии: инициации, элонгации и терминации. Для инициации транскрипции необходимо наличие специального участка в ДНК, называемого промотором. Когда РНК-полимераза связывается с промотором, происходит локальное расплетание двойной спирали ДНК и образуется открытый промоторный участок. Элонгация (удлинение) цепи РНК — это стадия транскрипции, которая наступает после присоединения 8 рибонуклеотидов. При этом движущаяся РНК-полимераза вдоль цепи ДНК действует подобно застежке молнии, раскрывая двойную спираль, которая замыкается позади фермента по мере того, как соответствующие основания РНК спариваются с основаниями ДНК. Терминация (прекращение роста) цепи мРНК происходит на специфических участках ДНК, называемых терминаторами. Для эукариот имеется ряд принципиальных отличий в этих процессах. Для эукариот характерно «кэпирование» 5—конца мРНК (первыми включаются нуклеотиды, содержащие основания аденин или гуанин). Модификация 5— конца мРНК приводит к образованию особой последовательности РНК, называемой кэп-структуры. При модификации 3—конца к нему присоединяются последовательность poly(A) длиной 20-200 нуклеотидов (poly-A — «хвост»). Первичным продуктом гена, формируемым в ядре, состоящем из больших по размерам молекул-предшественников, называемым незрелой, или гетерогенной ядерной РНК (гяРНК), является точная копия транскрибированного участка ДНК. Процесс формирования зрелых молекул РНК из предшественников называется процессингом, в результате которого молекулы подвергаются модификации по 5' --> 3' концам и сплайсингу. Сплайсинг гетерогенной ядерной РНК — это удаление последовательностей РНК, соответствующих нитронам ДНК, и соединение участков с транскрибируемыми последовательностями экзонов (рис. 3.5) (см. гл. 4.2).  Для эукариот характерно большое число реакций сплайсинга. Это происходит благодаря наличию в интронах строго определенной последовательности оснований ГУ или ГА в начале соответствующего участка интрона РНК и оснований АГ — в конце. Молекулы гяРНК имеют молекулярную массу более 107, в то время как молекулярная масса мРНК не превышает 2 х106. Трансляция (перевод) — процесс воплощения генетической информации мРНК в структуру полипептида. Это второй этап белкового синтеза, осуществляемый последовательной поликонденсацией отдельных аминокислотных остатков, начиная с аминоконца полипептидной цепи к карбоксильному концу. Зрелая матричная РНК выходит в цитоплазму, где осуществляется процесс трансляции — декодирование мРНК в аминокислотную последовательность белка. Процесс декодирования осуществляется в направлении от 5' —» 3' и происходит в рибосомах. Комплекс мРНК и рибосом называется полисомой. Подобно транскрипции механизм трансляции состоит из трех этапов: инициации, элонгации и терминации. Трансляция начинается со стартового кодона АУГ, который при локализации в смысловой части структурного гена кодирует аминокислоту метионин. Каждую аминокислоту доставляет к полисоме транспортная РНК (тРНК), специфичная к данной аминокислоте. тРНК выполняет роль посредника между кодоном мРНК и аминокислотой. Молекулы тРНК узнают в цитоплазме соответствующий триплет (кодон в мРНК) по принципу спаривания комплементарных азотистых оснований. тРНК, которая подходит к малой субчастице, образует связь кодон — антикодон, при этом одновременно передает свою аминокислоту в ами-ноацильный участок (А-участок) большой субъединице. К кодону АУГ «подходит» антикодон только той тРНК, которая переносит метионин. Поэтому прежде всего к рибосоме доставляется метионин. Затем кодон АУГ переходит на пептидиль-ный участок большой субъединицы (Р-участок). В результате этих процессов образуется транслирующая рибосома — инициирующий комплекс. Элонгация — это последовательное включение аминокислотных остатков в состав растущей полипептидной цепи. Каждый акт элонгации состоит из трех этапов: О узнавание кодона, которое заключается в связывании антикодона с очередной молекулой тРНК, несущей аминокислоту, с кодоном свободного А-участка на рибосоме; О образование пептидной связи, которое происходит лишь тогда, когда оба участка А и Р заняты молекулами тРНК. Часть большой субъединицы рибосомы — фермент пептидилтрансферазу, катализирующий образование пептидной связи; О транслокация, где тРНК участка Р, не связанная с пептидом, покидает рибосому. Затем молекула тРНК с полипептидом переходят из А на Р-участок и, наконец, рибосома перемещается вдоль РНК на один кодон. Терминация (окончание синтеза) происходит по команде кодонов УАА, УАГ, УГА. В природе не существует таких молекул тРНК, антикодоны которых соответствовали бы этим кодонам. Каждая мРНК транслируется, как правило, несколько раз, после чего разрушается. Среднее время жизни молекулы мРНК около 2 мин. Разрушая старые и образуя новые мРНК, клетка может довольно строго регулировать как тип продуцируемых белков, так и их количество. Это регуляция синтеза белка на уровне транскрипции. У эукари-от возможна регуляция и на уровне трансляции. Синтез белка — один из существеннейших показателей жизни. Жизнь каждого индивидуума начинается с оплодотворенной яйцеклетки, которая многократно делится. Вскоре в образовавшейся клеточной массе начинается дифференци-ровка: между многими ранее однородными клетками возникают различия. Клетки дифференцируются потому, что в них содержатся разные белки, от присутствия которых зависит, какие реакции будут проистекать в клетке, а также свойства и функции данной клетки. Поскольку любой белок является продуктом гена, то дифференци-ровка обусловлена тем, что разные гены включаются и выключаются на разных этапах онтогенеза. Кроме того, каждый человек на Земле в прошлом, настоящем или будущем имеет свой неповторимый набор только ему свойственных белков, именно поэтому каждый человек уникален. Исключение составляют монозиготные близнецы, у которых генетический материал идентичен. Именно специфичность белковых наборов обеспечивает иммунный статус каждого человека. 3.4. Дополнение. Полимеразная цепная реакция Полимеразная цепная реакция (ПЦР) была открыта в 1984 г. Кэри Б.Мюллисом. Она основана на том, что новосинтезируемые цепи нуклеиновых кислот могут служить матрицами в следующих циклах репликации. Принцип метода ПЦР показан на рис. 3.6. ПЦР осуществляется последовательными циклами. В каждом цикле происходят следующие события: 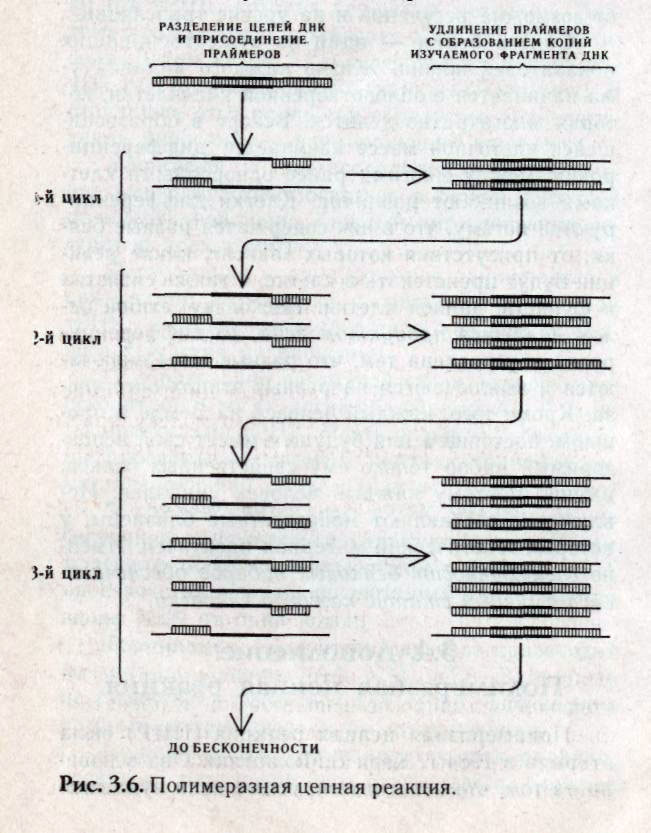 двухспиральная ДНК при нагревании разделяется на составляющие одноцепочечные цепи и в таком состоянии может служить матрицей для репликации; далее одноцепочечные нити ДНК инкубируют в присутствии ДНК-полимеразы и раствора, содержащего смесь всех четырех нуклеотидов, а также специфические последовательности ДНК (праймеры), что приводит к синтезу копий двух молекул ДНК. Затем процедуры повторяются сначала, и происходит копирование как старых, так и новых од-ноцепочечных цепей с образованием третьей и четвертой копий молекулы ДНК, затем все четыре снова копируются, и образуется уже восемь молекул ДНК, и т.д. число растет в геометрической прогрессии. В результате 20-30 циклов амплифицируется (увеличивается число копий) эффективное количество ДНК- Отдельный цикл занимает около 5 мин, а для бесклеточного молекулярного клонирования фрагмента ДНК требуется всего несколько часов. Метод ПЦР отличается очень высокой чувствительностью: он позволяет обнаружить в пробе всего одну присутствующую в ней молекулу ДНК. Тот же способ пригоден и для анализа следовых последовательностей РНК, для этого РНК переводят в последовательности комплементарной ДНК (кДНК), используя обратную транскриптазу. Метод получил широкое использование в пренатальной диагностике наследственных болезней, выявлении генетическую «дактилоскопию» даже по одной единственной клетке (см. раздел 4.3). 3.5. Задание 1. Какова молекулярная масса гена двух цепей ДНК, если в одной цепи запрограммирован белок с молекулярной массой 1500? Примечание: молекулярная масса одной аминокислоты в среднем — 100, одного нуклеотида — 345. 2. Исследования показали, что 34% общего числа нуклеотидов мРНК приходятся на гуанин, 18% на цитозин. Определите процентный состав азотистых оснований, соответствующих двухцепо-чечной ДНК. 3. На фрагменте одной цепи ДНК нуклеотиды располагаются в последовательности ТТЦТЦТАЦ-ГТАТ. Нарисуйте схему двухцепочечной молекулы ДНК, объясните, каким свойством ДНК при этом вы руководствовались? Какова длина в нм, если каждый нуклеотид занимает 0,34 нм по длине цепи ДНК- Сколько содержится нуклеотидов в этой последовательности ДНК? 4. Участок полипептида представлен следующими аминокислотами: -сер-вал-глут-мет-тир-ала-вал-. Какое количество нуклеотидов входит в состав гена? 5. Какую длину имеет участок молекулы ДНК, кодирующий участок полипептида, содержащего 20 аминокислот, если расстояние между двумя нуклеотидами равно 0,34 нм? 6. В чем выражается свойство триплетности генетического кода (выберите одно из утвержде- ний): в кодировании трех аминокислот одним нук-леотидом; в соответствии одной аминокислоте трех триплетов нуклеотидов? 7. В чем выражаются свойства универсальности генетического кода (выберите одно из утверждений): в соответствии одних и тех же триплетов ДНК одним и тем же аминокислотам; в соответствии нескольких триплетов одной аминокислоте; генетический код един для всех организмов; в соответствии аминокислотам определенных триплетов? 8. Что такое вырожденность генетического кода (выберите одно из утверждений): аминокислоты кодируются несколькими триплетами; один и тот же триплет кодирует несколько аминокислот и большинство аминокислот кодируется несколькими триплетами? 9. Рибонуклеаза поджелудочной железы содержит в кодирующем участке ДНК 42 нуклеоти-да. Укажите количество аминокислот, входящих в этот белок. 10. Молекула инсулина состоит из 51 аминокислотного остатка. Сколько нуклеотидов несет участок ДНК, кодирующий данный белок? 11. Дан участок ДНК — ГТТЦТААААГГГЦЦЦ... Какова структура закодированного гена? Какова структура белка, если под воздействием химических мутагенов восьмой нуклеотид будет заменен цитозиновым? К каким биологическим последствиям это может привести? Перейдет ли такое изменение ДНК к потомству? 12. Последовательность нуклеотидов на мРНК ЦГГГГЦУУЦУАГААЦГАУГАГ. Укажите соответствующий этой последовательности участок гена антисмысловой нити ДНК, а также фрагмент белка, соответствующий данному участку ДНК. вирусных инфекций, а также в судебной медицине, поскольку дает возможность проводить 13. Какие изменения произойдут в строении белка, если на участке гена ТААТЦАААГААЦАААА между 10 и 11 нуклеотидами включается цитозин, между 13 и 14 тимин, и в конце добавляется аденин? 14. Участок гена, кодирующий полипептид, имеет следующий порядок азотистых оснований: ААААЦЦААААТАЦТТГТАЦГА. Во время репликации четвертый слева аденин выпал. Определите структуру полипептидной цепи в норме и после выпадения аденина. 15. Всякое ли изменение последовательности нуклеотидов ДНК сказывается на структуре и функции белка? Ответ поясните. 16. Сколько белков может кодировать один и тот же участок ДНК? |