УЧЕБНИКгенетика. Генетика изучает процессы преемственности жизни на молекулярном, клеточном, организменном и популяционном уровнях
 Скачать 6.93 Mb. Скачать 6.93 Mb.
|

|
Глава 10 ГЕНЕТИКА ПОЛА 10.1. Хромосомный механизм определения пола В кариотипе человека из 46 хромосом 44 одинаковы у всех особей, независимо от пола (эти хромосомы называют аутосомами), а одной парой хромосом, называемых половыми, женщины отличаются от мужчин. Это общебиологическая закономерность для всех живых организмов, размножающихся половым путем. У женщин 2 половые хромосомы одинаковы (гомологичны), их называют Х-хромосомами. У мужчин пара половых хромосом представлена гетерохромосомами, так как они неодинаковы: одна из них Х-хромосома (т.е. такая же, как у женщин), другая У-хромосома. В основе определения пола у человека лежит хромосомный механизм, реализующийся в момент оплодотворения. Поскольку у женщин половые хромосомы одинаковы, то каждая яйцеклетка несет Х-хромосому, такой пол называют гомогаметным. У мужчин в процессе гаметогенеза формируется два типа гамет в равной пропорции: Х-сперматозоиды и У-сперматозоиды. Это биологическая закономерность, обусловленная механизмом мейоза. Мужской пол называется гетерогаметным. Хочется отметить, что теоретически соотношение полов должно быть 1:1 Это статистическая закономерность, обеспечиваемая условием равновероятной встречи гамет. Пол будущего потомка всегда определяет гетерогаметный пол (т.е. мужской). При патологии нерасхождения половых хромосом в гаметогенезе решающим фактором в определении пола у человека является наличие У-хромосомы или ее фрагмента. В таких случаях при любом числе Х-хромосом будет формироваться мужской пол. В случае отсутствия У-хромосомы или ее фрагмента будет формироваться женский пол (табл. 10.1). Таблица 10.1 Хромосомный механизм определения пола у человека в норме и при нерасхождении Х-хромосом
В настоящее время принято различать следующие уровни половой дифференцировки: 1. Хромосомное определение пола — 46, XX или 46, ХУ. 2. Определение пола на уровне гонад (яичники, или семенники). 3. Фенотипическое определение пола (мужчина или женщина, формирование вторичных половых признаков 4. Психологическое определение пола. 5. Социальное становление пола. Анализ нарушений числа и структуры половых хромосом позволил понять не только хромосомный механизм определения пола, но и получить информацию о гонадном и фенотипическом уровнях становления. Было показано, что инициация роста и созревание тестикул, их дифференцировка и сперматогенез связаны с эухроматиновым районом У-хромосомы (Эйчвальд и Силмсер, 1955), контролирующим трансплантационный антиген (Н-У антиген). Миграция первичных клеток зародышевого пути в гонады не зависит от пола. В норме направление развития определяется наличием У-хромосомы (мужской пол) или ее отсутствием (женский пол). Это развитие зависит от Н-У антигена. В настоящее время существует гипотеза, подтвержденная экспериментальными данными (Ohno, 1976), о том, что Н-У рецепторы имеются на поверхности клеток гонад обоих типов. Совместная инкубация с Н-У антигеном индуцирует семенники, но если активность Н-У антигена подавлена, то индуцируются яичники. Предполагается, что Н-У антиген кодируется не У-хромосомой, как думали ранее, а структурным аутосомным геном, находящимся под контролем У-хромосомы. У всех организмов, не имеющих У-хромосомы, этот структурный аутосомный ген не активируется. Экспрессия этого гена индуцируется факторами, которые в норме определяются У-хромосомой. Следовательно, возможны мутации, при которых будет синтезироваться Н-У антиген, даже в случаях, когда клетки лишены У-хромосомы. Наблюдения показали, что для превращения зачатка в семенники необходима определенная минимальная концентрация Н-У антигенов. Развитие вторичных половых признаков обусловлено дифференцировкой гонад. Человек по своей природе бисексуален. Половые органы формируются из мюллеровых и вольфовых каналов. У женщин мюллеровы протоки развиваются в фаллопиевы трубы и матку, а вольфовы — атрофируются. У мужчин вольфовы каналы развиваются в семенные протоки и семенные пузырьки. Под влиянием хорионического гонадотропина матери в эмбриональных семенниках клетки Лейдига синтезируют стероидные гормоны (тестостерон). В клетках Сертоли синтезируется гормон, называемый мюллеровым ингибирующим фактором (MIF). Эти гормоны действуют на зачатки внешних и внутренних половых органов. Нормальные особи мужского пола развиваются только в случае, если все элементы «срабатывают» в определенное время в заданном месте (рис. 10.1). Незначительные отклонения в работе на различных уровнях становления вторичных половых признаков приводят к неполному развитию мужского фенотипа в организме с мужским генотипом (мужской псевдогермафродитизм). При полном отсутствии всех элементов становления мужского пола формируются женские половые признаки, следовательно, становление женских половых признаков не нуждается в специальных регуляторных механизмах и является «конститутивным». В связи с этим Джост писал: «Становление мужского организма — это длительное, нелегкое и рискованное предприятие, своего рода борьба против имманентного стремления к женственности». 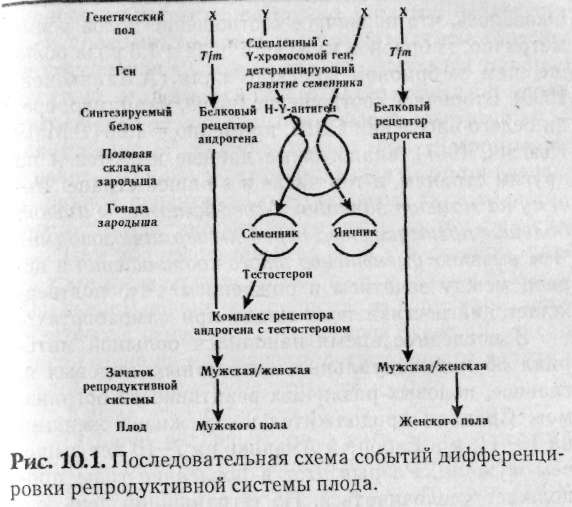 Описано не менее 19 различных дефектов генов как сцепленных с Х-хромосомой, так и аутосомно-рецессивных, приводящих к нарушению дифференцировки внешних и внутренних половых признаков: нарушения синтеза андрогенов и хорионического гонадотропина, отсутствие рецепторов на клетках Лейдига, дефекты ферментов, участвующих в синтезе тестостерона, нечувствительность клеток вольфовых протоков или мочеполового синуса к тестостерону и т.д. (Фогель и Мотульски, 1990). 10.2. Соотношение полов Различают первичное соотношение полов, точнее соотношение эмбрионов и вторичное — соотношение мальчиков и девочек среди новорожденных. Оказалось, что первичное соотношение полов асимметрично: эмбрионов мужского пола в 1,5 раза больше, чем эмбрионов женского пола (А.Балахонов, 1990). Вторичное соотношение полов, например, среди белого населения США, примерно — 1,06 (В.Мак-Кьюсик, 1967), аналогичные данные имеются и по другим странам, в том числе и в нашей стране. Почему на момент зачатия У-сперматозоиды имеют большие преимущества перед Х-сперматозоидами? Чем вызвано уменьшение этого соотношения в период между зачатием и рождением? Что подтверждает диагностика пола плода при самоабортах? В последнее время накопился большой материал об индивидуальных, возрастных, видовых и, главное, половых различиях реактивности организмов. Средняя продолжительность жизни женщин на 10—11, а в Европе и Америке на 7—10 лет выше, чем мужчин. Разрыв этот в последние годы продолжает увеличиваться. На сегодняшний день эти факты интерпретируют следующим образом. Мальчиков на момент зачатия значительно больше, так как Х-сперматозоиды значительно тяжелее, чем У-сперматозоиды (Х-хромосома значительно больше по размерам и относится к группе крупных хромосом, а У-хромосома — самая малая в кариотипе), в связи с чем У-сперматозоид, имея меньшую массу, имеет большую скорость передвижения и скорее достигнет цели, чем Х-сперматозоид. Низкая жизнеспособность мужских эмбрионов и в последующем — особей мужского пола объясняется другой гипотезой, связанной с уровнем метаболических процессов в организме, формирующем быстрый тип старения (короткожительство) и медленный тип старения (долгожительство). Мужской пол физически сильнее женского, но генетически слабее, что связано, возможно, с разной биологической значимостью полов. Считают (статистические исследования ВОЗ в 10 странах Западной Европы), что тип старения связан с Х-хромосомами: долгожительство — доминантно, а короткожительство — рецессивно. У мужчин только одна Х-хромосома, значит, если она может обеспечивать альтернативно с равной вероятностью доминантность и рецессивность, то генетическая вероятность долгожительства — 0,5 и короткожительства — 0,5. У женщин пара Х-хромосом, здесь вступают в силу законы Менделя (см.выше), значит, соотношение в популяции доминантных и рецессивных фенотипов будет 3:1, то есть среди женщин вероятность встречаемости долгожителей — 0,75 и короткожителей — 0,25.
10.3. Наследование признаков, сцепленных с полом Известно, что Х-хромосомы генетически активны, как любая из аутосом. У-хромосомы — генетически инертны, у человека лишь некоторые гены, не являющиеся жизненно важными, локализуются в ней (гипертрихоз — волосатые уши, некоторые формы аллергий, диспепсии), такие признаки передаются только от отца к сыну. Такой тип наследования по мужской линии называют голандрический (рис.10.2). 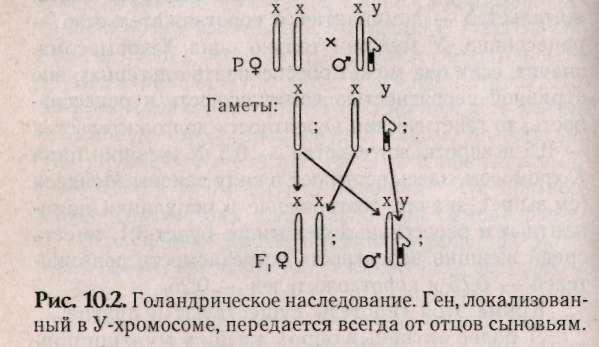 Законы передачи признаков, сцепленных с X-хромосомами, были впервые изучены Т.Морганом на дрозофилах. Кариотип самца дрозофилы несет пару различных хромосом, получивших название половых (ХУ), а самки — пару одинаковых половых хромосом — XX. Самки образуют только один тип гамет — X, а самцы два типа гамет Х- и У. Ген, определяющий белый цвет глаз, находится у дрозофил в Х-хромосоме (рецессивный аллель), доминантный — формирует красные глаза. При скрещивании мух с белыми глазами (white) — «w» и мух с обычными темно-красными глазами — W+ были обнаружены различия результатов реципрокных скрещиваний. Два скрещивания, которые характеризуются взаимно противоположным сочетанием анализируемого признака и пола, называются реципрокными скрещиваниями:  При скрещивании красноглазой самки и белоглазого самца (гомогаметный пол доминантен) в F, все мухи были красноглазые, а в F2 происходило расщепление 3 красноглазых : 1 белоглазой. Значит, если гомогаметный пол доминантен, то соблюдаются все законы Менделя: единообразие первого поколения и расщепление во втором поколении по фенотипу 3:1. При реципрокном скрещивании, когда гомогаметный пол был рецессивен (белоглазая самка и красноглазый самец), в первом поколении наблюдались иные результаты: все самки в F, были красноглазыми, а все самцы — белоглазыми. Такое наследование получило название крисс-кросс (или крест-накрест) наследования: сыновья наследуют фенотипический признак матери, а дочери — признак отца. Полученные результаты Морган объяснил тем, что ген «w» находит+ся в Х-хромосоме, а У-хромосома генетически инертна или, по крайней мере, не содержит гена «w». Этот тип наследования получил название наследования, сцепленного с полом. Таким образом, ген «w» сцеплен с полом, т.е. находится в Х-хромосоме. Установленные Т.Морганом законы справедливы и для человека. Проиллюстрируем законы наследования признаков, сцепленных с полом, установленные Морганом, на следующем примере. В брак вступают женщина-дальтоник (рецессивный признак) и мужчина с нормальным цветовосприятием (рис.10.3). Изучая цитологическую схему данного брака, видим, что сыновья единственную Х-хромосому получают от матери, следовательно, будут иметь ее фенотип по данному признаку (дальтоники). Дочери получают одну Х-хромосому от матери (несущую рецессивный аллель дальтонизма), а другую Х-хромосому от отца (несущую доминантный аллель цветовосприятия) и будут иметь нормальное зрение. Следовательно, фенотипический признак отца перешел к дочерям, а от матерей — к сыновьям (крисс-кросс наследование). 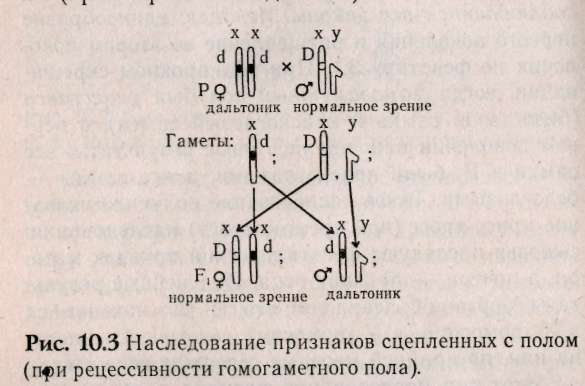 В случае, если мать имеет нормальное зрение, а отец дальтоник, все дети будут иметь нормальное цветовосприятие (рис. 10.4а), а если дочь с нормальным цветовосприятием, но носитель гена дальтонизма, выйдет замуж за здорового мужчину, то возможное соотношение фенотипов у детей будет 3 ; 1 (рис. 10.46). Сотрудник Т.Моргана КБриджес обратил внимание на редкие отклонения от схемы крисс-кросс наследования. Бриджес предположил, что такие отклонения в наследовании могут быть связаны с нарушением расхождения хромосом в мейозе. Он исследовал хромосомы таких исключительных мух и убедился, что белоглазые самки имеют У-хромосому наряду с двумя XX, а красноглазые самцы — одну Х-хромосому. Тем самым было впервые доказано, что определенный ген w находится в конкретной хромосоме — X. 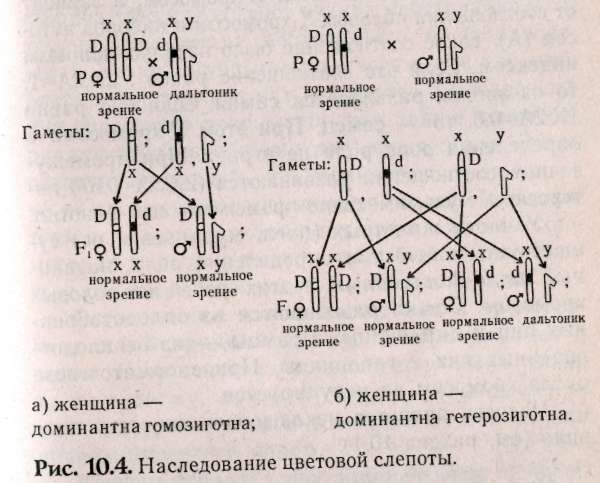 В F, от скрещивания красноглазой самки и белоглазого самца изредка встречались мухи, у которых один глаз белый, а другой красный. При более внимательном рассмотрении оказывается, что эти мухи симметрично представлены женской и мужской половинками тела. Таких мух называют билатеральными гинандроморфами. При этом белый глаз находится на мужской половине. Эти особивозникают в результате потери одной Х-хромосомы при первом дроблении зиготы, которая должна дать начало самке. Потери хромосом могут происходить и на более поздних стадиях развития. Тогда появляются организмы-мзаики, у которых в разных пропорциях представлены участки тела, состоящие из клеток с неодинаковым числом хромосом. Бриджес в 1922 г. показал, что пол у дрозофилы определяется не числом Х-хромосом, а зависит от соотношения половых Х-хромосом и наборов ауто-сом (А). Такое соотношение было названо половым индексом. Если это соотношение равно : 2Х:2А=1, то из зиготы развивается самка, если оно равно 1Х:2А=0,5, то — самец. При этом У-хромосома в определении пола роли не играет. При промежуточном соотношении развиваются (2Х:ЗА=0,67) ин-терсексы, мухи, имеющие промежуточный фенотип. У части животных (пчел, муравьев и ос) существует особый тип определения пола, названный гапло-диплоидным. У этих особей нет половых хромосом. Самки развиваются из оплодотворенных яиц и диплоидны, а самцы — из неоплодот-воренных яиц и гаплоидны. При сперматогенезе число хромосом не редуцируется. Человек является исключением из этого правила (см. раздел 10.1). 10.4. Наследование, ограниченное и контролируемое полом Гены, имеющиеся в кариотипе обоих полов, но проявляющиеся преимущественно лишь у одного пола, называются ограниченные полом. Эти гены могут быть не сцеплены с половыми хромосомами и локализованы в любой аутосомной хромосоме. Вот пример строгой ограниченности полом. Если S и s представляют собой два аллеля, выражение которых ограничено полом, то три генотипа SS; Ss; ss оказываются неразличимыми у одного пола, но дают два или три различных фенотипа у другого пола в зависимости от того, сходна ли гетерози-гота с одной из гомозигот или отличается от обеих. Различные анатомические и физиологические черты, присущие женскому полу, такие, например, как ширина таза или возраст начала менструации, контролируются генами, получаемыми от обоих родителей. Такие сугубо мужские черты, как характер роста волос на лице или количество и распределение волосяного покрова на теле также контролируются генами, общими для обоих полов. Здесь следует подчеркнуть, что ограниченность полом — не то же самое, что сцепление с полом. Последний термин касается локализации генов в половых хромосомах; первый термин — проявления генов только у одного из двух полов. Ограниченное полом наследование представляет собой крайний пример контролируемости полом. Если генотип проявляется у обоих полов, но по-разному, принято говорить о проявлении гена, контролируемого (зависимого) полом, или модифицируемого полом. Контролируемая полом доминантность, по-видимому, лежит в основе характера облысения. Изучение этой черты представляет существенные трудности в связи с тем, что фенотип облысения различно проявляется. Облысение может быть слабым или полным, лысина может локализоваться по-разному, проявляться в раннем или пожилом возрасте, иногда облысение связано с нарушением функции щитовидной железы или последствиями инфекционных болезней. У большинства — наследственная этиология. Признак может проявляться у обоих полов, но чаще и выраженнее проявляется у мужчин. Основоположником изучения признаков, контролируемых полом, является Бернштейн, который проанализировал наследование певческих голосов у взрослых европейцев. Его вывод — шесть различных певческих голосов (бас, баритон, тенор, сопрано, меццо-сопрано и альт) контролируются одной парой аллелей. Более поздние исследования показали, что тип голоса контролируется половой конституцией, начиная с периода полового созревания находится под влиянием половых гормонов. 10.5. Дополнение. Возможно ли предопределить пол у человека? В прошлом веке индейцы племени хавасупаи обладали удивительной способностью влиять на пол своих будущих детей. Сходный эффект неслучайного распределения полов известен у филиппинцев, индейцев Венесуэлы, аборигенов Австралии. Они не испытывали влияния цивилизации и практически не пользовались контрацепцией. Во Франции и Англии сразу после каждой из двух мировых войн наблюдалось странное возрастание числа мальчиков из числа новорожденных, отцы которых были солдатами. Отмечено, что профессия родителей или их заболевания могут влиять на пол детей. Например, среди детей водолазов, анестезиологов, пилотов и священников чаще встречаются девочки. Как и у мужчин, больных лимфо-мой Ходжкинса. У женщин с гепатитом А или шизофренией чаще рождаются дочери, чем сыновья. Существуют несколько гипотез, объясняющих, как происходит предопределение пола у человека. Гипотеза Мартина: среди сперматозоидов выделяют «зайцев» — У-сперматозоиды и «черепах» — Х-сперматозоиды. У-сперматозоиды активнее, чем Х-сперматозоиды, но быстрее погибают. Если яйцеклетка готова к оплодотворению, то первыми ее оплодотворяют У-сперматозоиды, а если нет, то вероятность оплодотворения Х-сперматозоидом возрастает, т.к. У-сперматозоиды быстро погибают. У молодоженов, часто занимающихся сексом, У-сперматозоиды постоянно присутствуют в половых путях женщины и оплодотворяют яйцеклетку. Спустя несколько лет ситуация меняется (когда секс становится эпизодическим) и яйцеклетки оплодотворяются более долгоживущими Х-сперматозоидами. Поэтому первые дети после свадьбы — обычно мальчики, а более поздние — девочки. Аналогично в ситуациях в семьях вернувшихся с фронта солдат. Другая гипотеза была предложена Джеймсом (Лондонский университетский колледж), утверждавшим, что закономерности распределения полов обусловлены гормональными изменениями в организме родителей. Он считает, что повышение уровня тестостерона и эстерогенов у обоих родителей увеличивает вероятность рождения мальчиков, а возрастание уровня гонадотропина — девочек. Предложения основаны на клинических наблюдениях: лечение бесплодия гонадотропинами у женщин приводило к рождению дочерей, а у мужчин — сыновей. Джеймс, как и Марти, считает, что при быстром оплодотворении чаще рождаются мальчики, но связывает с соотношением половых гормонов в момент зачатия. В первой половине менструального цикла до момента готовности яйцеклетки к оплодотворению уровень тестостерона и эстрогенов высок, что приводит к рождению мальчиков. Далее по циклу возрастает уровень гонадот-ропинов, которые обуславливают зачатие девочек. Доминирование в поведении и агрессивность связывают с высоким уровнем тестостерона в крови. Предварительные данные говорят о положительной зависимости между высоким социальным статусом женщины и рождением у нее сыновей, хотя исследования по выявлению влияния уровня тестостерона на пол будущего ребенка не проводились. Третья гипотеза выдвинута Триверсом и Вил-лардом (Гарвард). Они предположили, что соотношение полов у млекопитающих объясняется адаптивными механизмами (особенно у полигамных, проявляющих заботу о потомстве). Если самка ослабленная, то пол скорее всего будет женским (будущая самка, даже не очень сильная, сможет найти себе супруга). Если же у матери прекрасное физическое здоровье, то, скорее всего, это будет самец (сильный самец сможет иметь много детенышей, а слабый у полигамных видов часто не имеет потомства).-Гипотеза была подтверждена при изучении паукообразных обезьян. В применении к человеческому обществу эта гипотеза позволяет найти определенную взаимосвязь между социальным статусом человека и полом его детей. Мюллер (Германия) собрал данные о статистически достоверном преобладании сыновей среди детей, рожденных в семьях с высоким социальным статусом. Обратная тенденция наблюдается среди людей низкого социального положения. Лоррен и Столковский (Франция) предложили метод, с помощью которого можно планировать пол будущего ребенка. Его суть в особом режиме питания, при котором в течение 6 недель, предшествующих зачатию, необходимо отдавать предпочтение определенным пищевым продуктам. Им была обследована экспериментальная группа женщин, из которых 87% родили ребенка запланированного пола. Оказалось, что женщины, которые хотят иметь сыновей, должны употреблять острые продукты с повышенным содержанием солей (натрия и калия). Женщины, желающие иметь дочерей, должны есть больше молочных продуктов, богатых кальцием и магнием. Женщины обязаны придерживаться установленного рациона. Те, кто уже имеет несколько дочерей, но хотят сына, должны выдержать рацион в течение 3-4 месяцев. Пока не ясно, каким образом рацион влияет на пол эмбриона. Возможно, в будущем человечество сумеет влиять на баланс своего потомства. 10.6. Задание 1. Мужчина-гемофилик женится на здоровой женщине. У них рождаются здоровые дети, вступающие в брак с лицами, не страдающими гемофилией. Обнаружится ли у внуков гемофилия, если да, то с какой вероятностью возможно появление больных внуков и внучек? 2. У человека в У-хромосоме локализован ген, определяющий развитие перепонок между вторым и третьим пальцами ног. Определите, какие будут дети и внуки в браке мужчины, имеющего этот признак, и здоровой женщины. 3. Ангидрозная эктодермальная дисплазия (отсутствие эмали на зубах) передается как рецессивный, сцепленный с Х-хромосомой признак. В браке здоровой женщины и мужчины, больного ангид-розной эктодермальной дисплазией, рождаются больная девочка и здоровый сын. Определите вероятность рождения следующего ребенка здоровым. 4. Одна из форм гемералопии (куриная слепота) наследуется как рецессивный, сцепленный с X-хромосомой признак. У здоровых родителей рождается сын с гемералопией. Оцените вероятность рождения в этой семье здоровых дочерей и сыновей. 5. В семье, где отец имел гипертрихоз (сцеплен с У-хромосомой), а мать полидактилию (аутосом-но-доминантный признак), родилась нормальная в отношении двух признаков дочь. Какова вероятность того, что следующий ребенок будет без аномалий, каков пол возможен у этого ребенка? 6. В браке здоровых родителей родился сын-альбинос (аутосомно-рецессионный признак), страдающий гемофилией. Определите вероятные генотипы родителей, а также оцените вероятность рождения второго сына здоровым? 7. Женщина, страдающая катарактой (доми-нантно-аутосомная форма), с нормальной свертываемостью крови выходит замуж за гемофилика, имеющего нормальное зрение. У них родился сын-гемофилик. Каков прогноз в отношении здоровья детей в этой семье? 8. Талассемия — неполностью доминантный аутосомный признак (гомозиготы почти всегда погибают). Дальтонизм — рецессивен, сцеплен с X-хромосомой. Дигетерозиготная женщина вступает в брак с мужчиной, страдающим легкой формой талассемии, но с нормальным зрением. Какова ве- роятность рождения полностью здоровых детей в этой семье? 9. Гипертрихоз сцеплен с У-хромосомой, а ихтиоз сцеплен с Х-хромосомой. В семье, где мать здорова по двум признакам, а мужчина — обладатель гипертрихоза, родился мальчик с признаком ихтиоза. Определите вероятность рождения здоровых детей. 10. В брак вступают родители с нормальной свертываемостью крови, имеющие II и III группы крови. У них рождается сын-гемофилик, имеющий I группу крови. Укажите генотипы родителей и генотипы детей, возможных в этой семье. 11. В предложенной табл. 10.2 по вертикали указаны генотипы и фенотипы женщин, а по горизонтали указаны только фенотипы мужчин. Таблица предполагает равновероятность возможных сочетаний генотипов мужчин и женщин. Все возможные потомки мужчины (от предполагаемых браков с перечисленными женщинами) указаны в столбце под его фенотипом. Необходимо определить генотип каждого из мужчин, анализируя приведенные соотношения фенотипов детей. Студенту необходимо выбрать правильный ответ возможного потомства для каждой из возможных пар и проставить цифры ответов (соответствующих номеру строки, где они стоят) в месте пересечения фенотипов родителей. Например, в браке голубоглазых родителей-дальтоников все возможные дети будут голубоглазыми дальтониками (правильный ответ надо искать во втором столбце — под фенотипом мужчины, он расположен в третьей строке сверху, цифру 3 надо проставить в эталоне для ответа на пересечении указанных фенотипов родителей). 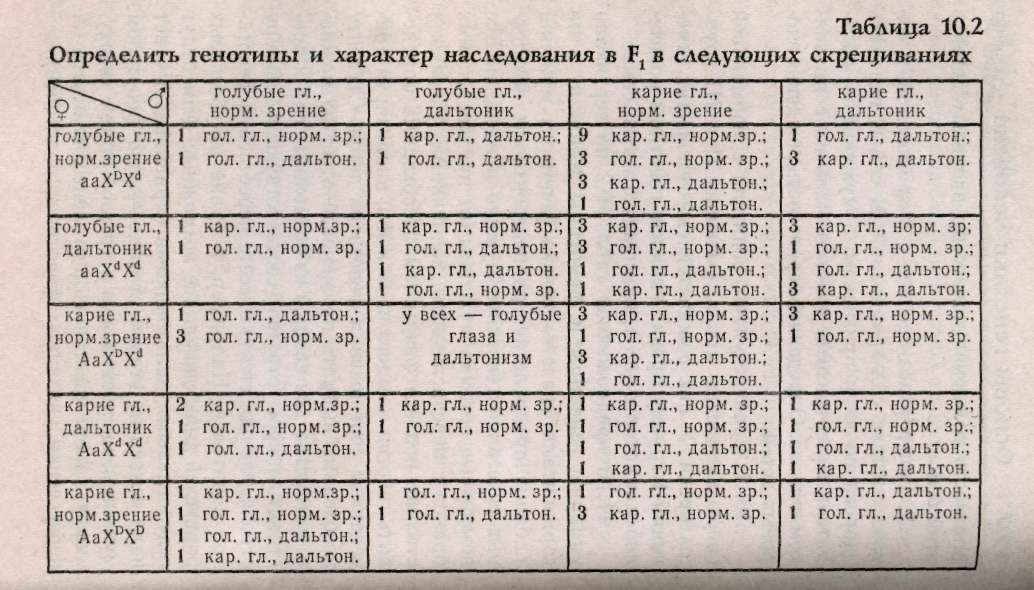 |