УЧЕБНИКгенетика. Генетика изучает процессы преемственности жизни на молекулярном, клеточном, организменном и популяционном уровнях
 Скачать 6.93 Mb. Скачать 6.93 Mb.
|

|
Глава 8 Взаимодействие генов 8.1.Комплементарность Например, чтобы человек имел нормальный слух, необходима согласованная деятельность нескольких пар генов, каждый из которых может быть представлен доминантным и рецессивным аллелями. У человека слух нормален, если каждый из них имеет хотя бы по одному доминантному аллелю в диплоидном наборе хромосом. В случае, если хотя бы один из них представлен гомозиготой рецессивной, человек будет глухим. Мы разберем это на простом примере, предположив., что нормальный слух формирует пара генов, и рассмотрим явление комплементарности. В брак вступает пара глухих, у них рождаются дети, которые все слышат. Это может быть только в случае, если один родитель страдает глухотой по гену «в» (ААвв), а другой — по гену «а» (ааВВ). Р ♀ ААвв х ♂ааВВ глухаяглухой Гаметы: Ав; аВ; F1 АаВв; слышащий Все дети получат с гаметами Ав и аВ доминантные аллели, их генотипы будут одинаковы — АаВв, то есть каждая гамета будет иметь доминантный аллель по каждому гену, которые будут взаимодействовать, дополняя друг друга и формировать новый, по отношению к родителям, признак — нормальный слух. Это и есть комплементарное взаимодействие генов. Допустим, что дети вступят в брак с себе подобными, и оценим вероятность рождения слышащих и глухих детей в этом случае. Р ♀АаВв х ♂АаВв слышащая слышащий
Таким образом, мы получили соотношение: 9 А-В- : 3 А-вв : 3 ааВ- : 1 аавв слышащие глухие глухие глухой или 9 слышащих : 7 глухих Соотношение фенотипических классов при комплементарном взаимодействии может быть разным, в зависимости от вида комплементарности и проявляемости отдельных генов: 9:3:3:1; 9:6:1; 9:7. 8.2. Эпистаз Эпистаз — подавление действия одного гена действием другого гена. Различают эпистаз доминантный (доминантный аллель эпистатирует, или подавляет проявление доминантного аллеля другого гена) и рецессивный (рецессивный аллель эпистатирует, ингибирует или подавляет проявление доминантного аллеля другого гена). Анализ при эпистатическом взаимодействии ведется так же на основе дигибридного скрещивания, как и в предыдущих случаях. Цифровые расщепления при эпистазе 12:3:1 (если каждый из двух генов имел самостоятельное проявление, при этом в первом поколении признак особей повторяет фенотип одного из родителей) и 13:3 (ген-ингибитор не имеет самостоятельного проявления, при этом фенотип родителей и первого поколения одинаков). Подавляющий ген называется геномсупрессором, или ингибитором, а подавляемый — гипостатическим. Этот тип взаимодействия наиболее характерен для генов, участвующих в регуляции работы генов в онтогенезе и работе иммунных систем человека (см. раздел 13.5). Известно немало примеров эпистатического взаимодействия локусов у человека, приводящих к тому, что тот или иной доминантный аллель у некоторых индивидуумов не получает фенотипи-ческого выражения. Подобным примером может служить полидактилия, которая наследуется, как правило, по доминантному типу. Иногда встречается у детей «совершенно здоровых» родителей. В этом случае предполагается, что действие данного аллеля у родителей подавлялось другими генами (см. раздел 11.4). 8.3. Полимерия Полимерия — обусловленность определенного (обычно количественного) признака несколькими эквивалентными (полимерными) генами. При таком взаимодействии индивидуальное проявление каждого усиливается в результате взаимодействия. Полимерия бывает двух видов' некумулятивная — когда не важно количество доминаитных генов в генотипе, а важно его присутствие; кумулятивная — когда число доминантных аллелей влияет на степень выраженности данного признака У человека по типу кумулятивной полимерии наследуется пигментация кожи: чем больше доминантных аллелей, тем больше меланина образуется и тем интенсивнее окраска кожи. Разберем следующий пример, в котором необходимо определить вероятность появления детей с разными фенотипами в браке мулатов (особи дигетерозиготны). 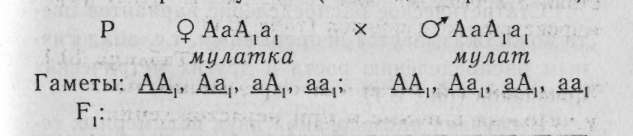
— негры (АААА) 1/16, — светлокожие негры (АААа) 4/16, — мулаты (ААаа) 6/16, — светлокожие мулаты (Аааа) 4/16, — белые (аааа) 1/16. Обратите внимание, что при полимерии гены, как правило, обозначаются одинаковыми буквами, чтобы подчеркнуть однонаправленность их действия, т.к. при формировании признака не важно, какому гену принадлежат доминантные гены, а важно их количество. Статистическое распределение вариантов цвета кожи оказывается непрерывным, т.е. аналогичным распределению роста и других метрических признаков (рис. 8.1). В случаях наследования двух полимерных генов цифровые расщепления при скрещивании гибридов будут иметь четкие зависимости, являющиеся модификациями все того же расщепления для двух генов 9:3:3:1. При кумулятивной полимерии соотношение фенотипов будет 1:4:6:4:1, а'при некумулятивной полимерии — 15:1. В представленном выше примере, если допустить, что нас интересует факт наличия пигментации у человека, а не ее интенсивности, получим соотношение, характерное для некумулятивной полимерии: 15 пигментированных к 1 белому. Если учесть, что спонтанные мутации происходят крайне редко (вероятность спонтанной мутации одного аллеля составляет 1/1000000-1/1000000000), то вероятность рождения у белых людей даже светлокожего мулата, а у негров — детей с более светлой кожей, крайне ничтожна. 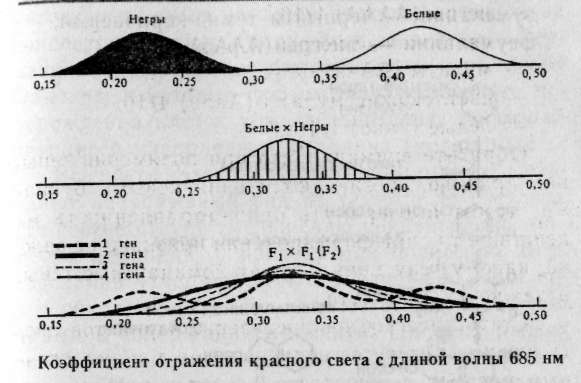 Рис. 8.1. Распределение цвета кожи у негров и людей белой расы. Цвет кожи оценивали по величине отражения света с длиной волны 685 нм. Кривые F2 представляют собой теоретически ожидаемые на основе различных предположений о числе генов, определяющих различие в цвете кожи между неграми и белыми. Исследование потомства F2 выявляет распределения, сходные с распределениями, полученными на основе предположения об участии трех или четырех пар генов [по W.F.Bodmer, L.L.Cavalli-Sforza (1976). Genetics, Evolution and Man, Freeman, San Francisco]. Меланин — пигмент, определяющий все мно-гобразие окрасок покровов, синтезируется в организме из аминокислоты тирозина в несколько этапов (рис. 8.2). Формирование промежуточного продукта диоксифениланина (ДОФА) — медиатора симпатической нервной системы свидетельствует о возможном плейотропном влиянии генов, участвующем в синтезе меланина, на свойства нервной системы. Меланин, формируя окраску покровов, является протектором от воздействия на организм УФ-лучей. Существуют два типа меланинов: эумеланин — черный и темно-коричневый; феумеланин — желтый и рыжий. 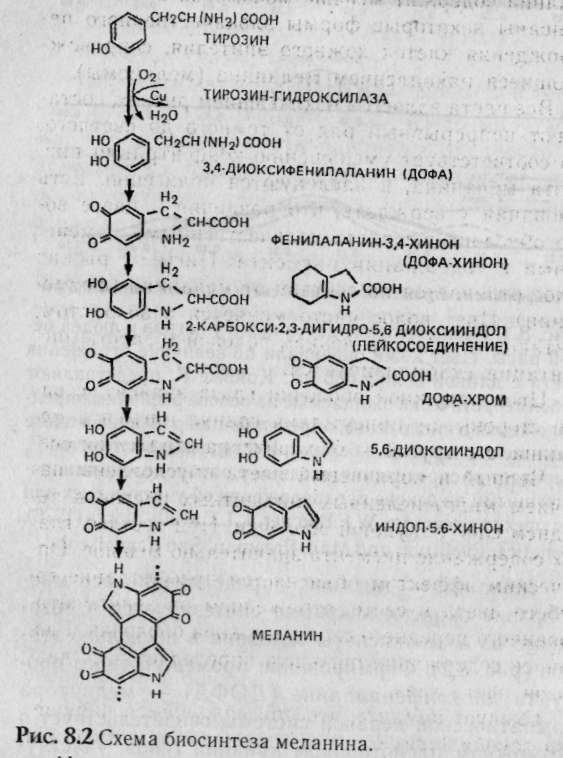 Показано, что на их синтез оказывает влияние метаболизм в клетках: при усиленном митозе клеток в основании волоса образуется феумеланин, а при замедлении эумеланин. При образовании фёумеланина важным условием является повышенная концентрация сульфгидрильных соединений, Феумеланин содержит меньше мономеров в молекуле. Описаны некоторые формы злокачественного перерождения клеток кожного эпителия, сопровождающиеся накоплением меланина (меланомы). Все цвета волос, за исключением рыжих, составляют непрерывный ряд от темного до светлого, что соответствует уменьшению концентрации пигмента меланина, и наследуются полигенно. Есть основания утверждать, что различия в цвете волос обусловлены чисто количественными изменениями в содержании пигмента. Пигмент рыжих волос отличается по составу от меланина (феумеланин). Цвет волос часто меняется с возрастом, считают, что с наступлением половой зрелости пигментация стабилизируется. Цвет радужной оболочки глаза зависит, с одной стороны, от присутствия гранул пигмента меланина, а с другой — от характера отражения света. Черный и коричневый цвета обусловлены наличием многочисленных пигментных клеток в переднем слое радужной оболочки. При светлых глазах содержание пигмента значительно меньше. Оптическим эффектом объясняется преобладание голубого цвета в свете, отраженном от депигменти-рованного переднего слоя радужной оболочки. Различное содержание пигмента определяет весь диапазон цвета глаз. Следует помнить, что при кажущемся нарушении закона Менделя о независимом наследовании признаков для двух генов, связанного с тем или иным видом взаимодействия генов, в основе «неожиданных» цифровых соотношений лежит классическое расщепление 9:3:3:1. Кроме того, в каждом конкретном случае необходимо понять, как происходит объединение расщепляющихся классов в единые фенотипы, только после этого можно интерпретировать результаты. Термин «взаимодействие генов» довольно условен, поскольку взаимодействуют не сами гены, а их продукты. 8.4. Гены-модификаторы Генами-модификаторами называют такие гены, которые регулируют активность других генов, усиливая или ослабляя их проявления. В таком случае получается, что в формировании признаков, кроме «основных» генов, принимают участие гены-модификаторы. Такой тип наследования встречается очень часто. Примером модифицирующего аллеля у человека может служить брахидактилия. Существуют различные формы проявления: резкое укорочение указательного пальца и незначительное укорочение. Большинство больных с брахидак-тилией гетерозиготны (ВЬ). Анализируя родословную, можно предположить наличие гомозиготного рецессивного аллеля n-модификатора, определяющего крайнее проявление гена, и доминантного аллеля N — определяющего слабое проявление. Исходя из этого, генетическая конституция у индивида с выраженной брахидактилией будет Bbnn, у индивидов со слабой брахидактилией BbNn либо BbNN, а нормальными могут быть индивиды, принадлежащие к трем различным генотипам: bbnn; bbNn; bbNN (Штерн, 1965). Гены-модификаторы у людей выявляются очень сложно. Поэтому признание их наличия не помогает предсказать специфический характер проявления признака в будущих поколениях. И все же важно знать о тех осложнениях, которые могут быть вызваны генами-модификаторами. Природа генов-модификаторов до сих пор не выяснена: то ли это истинные гены-модификаторы, влияющие на действие основных генов, или это эффект плейотропии. 8.5. Плейотропия Явления взаимодействия генов указывают на целостность генотипа при индивидуальном развитии особи, а признание принципа взаимодействия допускает, что все гены так или иначе взаимосвязаны в своем действии. Исходя из этого, мы можем сказать, что и один ген может влиять на несколько признаков. Такое множественное действие гена называют плейотропией. При плейотропии гены взаимодействуют на уровне продуктов контролируемых ими реакций, т.е. каждый ген ответствен за определенный этап метаболизма. Нарушения метаболизма на каком-либо из этапов отразятся на последующих (т.е. на нескольких элементарных признаках). У человека синдром Марфана (впервые описан в 1896 г.) обуславливается аутосомно-доминантным аллелем с высокой пенетрантностью и различной экспрессивностью. Синдром Марфана связан с высоким ростом, значительным удлинением костей пальцев ног и пальцев рук (паучьи пальцы — арахнодактилия), гиперподвижностью суставов, подвывихом хрусталика и аневризмом аорты. Соотношение полов равное, популяционная частота 0.04 на 1000 (Мак-Кьюсик). Такого рода аномалии скелета часто сопровождаются ненормальным расположением хрусталиков и пороками сердца. Разные симптомы, по Мак-Кьюсику, Нет стр 136-137 9:3:3:1; 1:4:6:4:1; 12:3:1; 13:3; 9:7; 9:3:4. • Какой тип взаимодействия генов имеет место в данной задаче — полимерия, эпистаз, компле-ментарность? • Какова вероятность рождения слышащих детей в этой семье? 1/16; 3/16; 9/16; 7/16; 13/16; 12/16; 1/4; 1/2. • Какое возможно потомство у этого ребенка, если он вступит в брак со здоровой женщиной, в семье которой никогда не было зарегистрировано случаев глухоты? Все дети будут здоровы; все дети больны; 25% здоровых детей; 50% здоровых детей; 12,5% здоровых детей. 8.7. Лабораторная работа. Создайте лицо Наверное, вы не раз удивлялись, почему все люди так не похожи друг на друга, даже близкие родственники не выглядят точной копией друг друга. Это связано с большим разнообразием существующих характерных черт в человеческой популяции и новыми их сочетаниями, образующимися в процессе репродукции человека. Причины того, почему братья и сестры имеют различные генотипы, могут быть исследованы в результате проведения данной работы. Возьмите монетку: орел — доминантный признак; решка — рецессивный. Подбрасывая монетку, определите, кто из родителей передал ребенку доминантный или рецессивный признак. Допустим, что ваш ребенок имеет гетерозиготу для данной черты лица, представленной в этой лабораторной работе. Заполните предлагаемую таблицу и нарисуйте портрет ребенка, который у вас в этой работе получится. 1. Определите пол ребенка. Генотип девочки XX (решка), генотип мальчика XY (орел). 2. Дайте имя ребенку. 3. Определите черты лица. 4. Нарисуйте портрет вашего ребенка и пусть ваш ребенок выглядит так, как если бы он (она) достиг(ла) совершеннолетия. 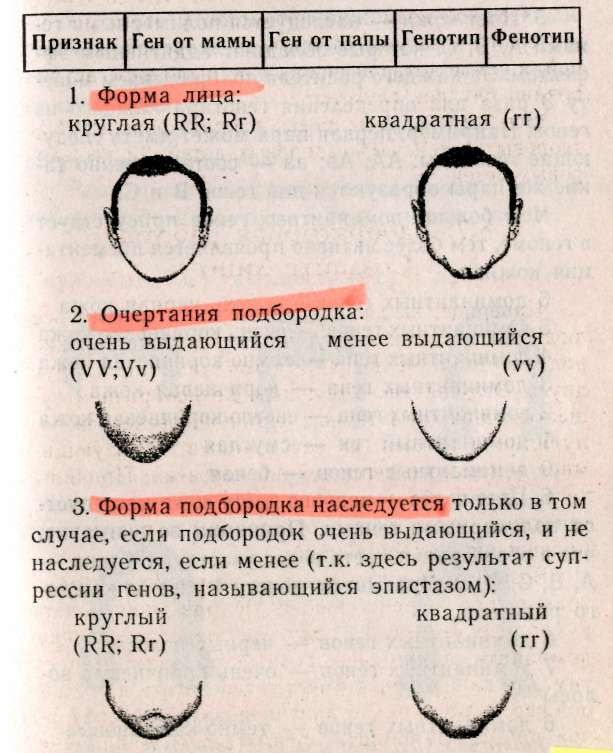  5. Цвет кожи — наследуется полимерными генами А, В, С, которые обладают аддитивным эффектом. За каждого родителя подбрасываем монету 3 раза для определения генотипа каждого из генов. Например, первая пара может иметь следующие генотипы: АА; Аа; аа — соответственно Такие же пары образуются для генов В и С. Чем больше доминантных генов присутствует в геноме, тем более активно проявляется пигментация кожи: 6 доминантных генов — очень черная кожа 5 доминантных генов — очень коричневая кожа 4 доминантных гена — темно-коричневая кожа 3 доминантных гена — коричневая кожа 2 доминантных гена — светло-коричневая кожа 1 доминантный ген — смуглая О доминантных генов — белая 6. Цвет волос — подобно цвету кожи наследуется полимерными генами. Принимая во внимание, что в образовании цвета волос участвуют 4 гена — А, В, С, D, подбрасываем монету 4 раза за каждого родителя. 8 доминантных генов — черные волосы 7 доминантных генов — очень коричневые волосы 6 доминантных генов — темно-каштановые 5 доминантных генов — каштановые 4 доминантных гена — светло-русые 3 доминантных гена — рыжеватая блондинка 2 доминантных гена — блондинка 1 доминантный ген — очень светлая блондинка О доминантных генов — белые (альбинос) 7. Рыжие волосы определяются единственным геном, представленным двумя аллелями Red (R) и red (г), и проявляются только в сочетании с доминантным геном, RR — ярко-рыжие; Rr — светло-рыжие; гт —отсутствие рыжих волос. Рыжие волосы проявляются только если у вашего ребенка меньше 6 доминантных генов. 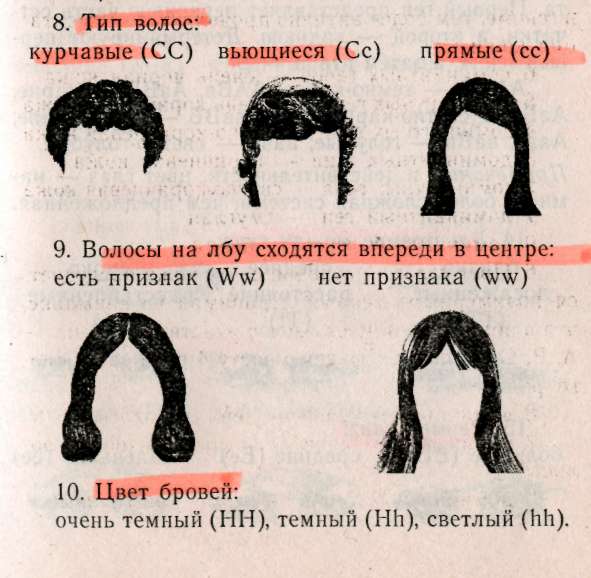 Нет стр.142-143 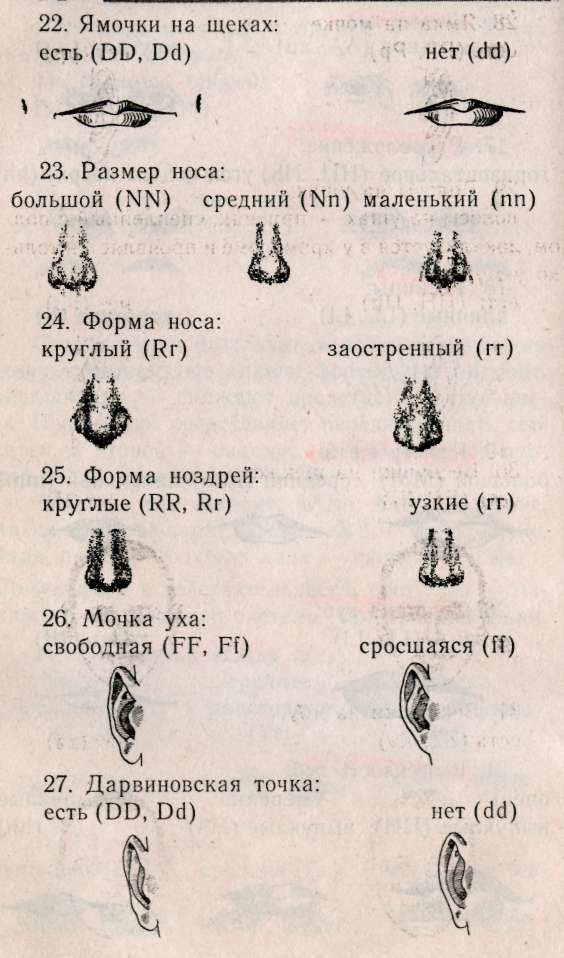 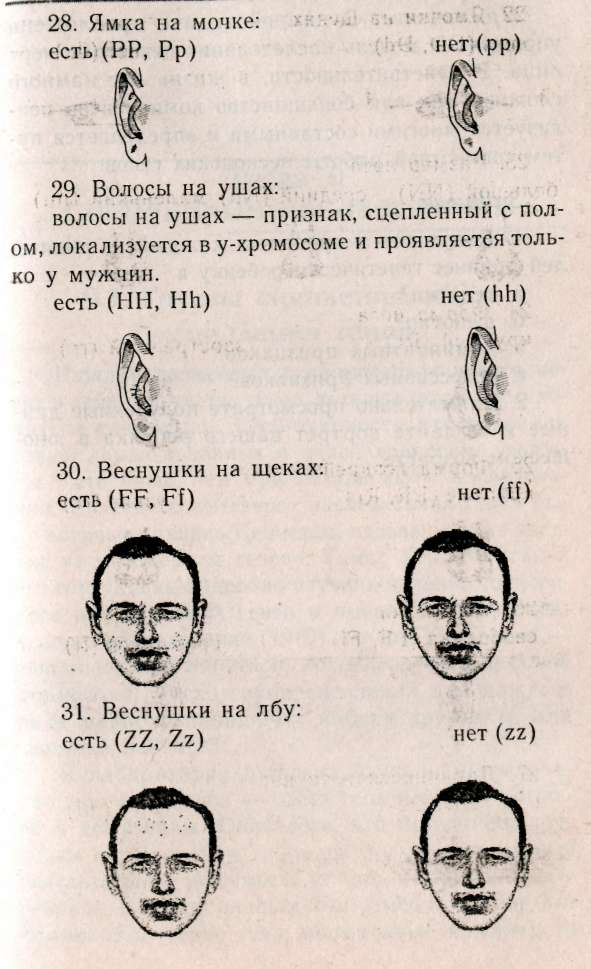 Примечание: В данной задаче предложена упрощенная модель наследования некоторых черт лица. В действительности, в жизни все намного сложнее, так как большинство компонентов реализуется многими составными и определяется путем совместной работы нескольких генов. Задание. 1. Как много в процентах каждый из родителей привнес генетически ребенку в а. генотип? б. фенотип? в. доминантных признаков? г. рецессивных признаков? 2. Внимательно просмотрите полученные данные и создайте портрет вашего ребенка в юношеском возрасте. Глава 9 СЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ 9.1. Группа сцепления, виды сцепления генов Каждая хромосома в кариотипе человека несет в себе множество генов, которые могут наследоваться совместно. Закономерности наследования генов, локализованных в одной хромосоме, должны быть иные, чем при независимом наследовании. Явление совместного наследования генов было впервые описано Пеннетом, назвавшим это явление «притяжением генов». Томас Гент Морган и его сотрудники подробно изучили явление сцепленного наследования генов и вывели законы сцепленного наследования (1910). Группа сцепления — это совокупность генов, локализованных в одной хромосоме. Число групп сцеплений для каждого вида равно гаплоидному набору хромосом, для человека равно 23. В лаборатории Моргана изучали наследование двух признаков — цвета тела и формы крыльев у дрозофилы. Оказалось, что при анализирующем скрещивании гибридов получались разные результаты в зависимости от того, какой пол был у гибрида. Если гибридным был самец (фенотип был доминантен: серое тело, нормальные крылья), то потомство в анализирующем скрещивании давало расщепление 1:1, т.е. получалось' 2 фенотипических класса, вместо ожидаемых по Менделю — четы рех. Причем эти два класса имели фенотипическое выражение родительских особей гибрида. Если гиб-' ридной была самка, то получались 4 фенотипических класса в неравных пропорциях: большую часть из них составляли особи, повторяющие фенотипы родителей гибрида, меньшую — особи с новыми сочетаемыми признаками. Сколько бы не повторялись опыты, результаты были неизменны. Постоянство результатов позволило сделать Моргану следующие выводы: сцепленное наследование может быть полным — явление полного сцепления генов, при котором группа сцепления не нарушается, все гены наследуются совместно (гибридный самец), и неполным (гибридная самка) — группа сцепления нарушается явлением кроссинговера. 9.2. Кроссинговер Кроссинговер — обмен идентичными участками между гомологичными хромосомами, приводящий к рекомбинации наследственных задатков и формированию новых сочетаний генов в группах сцепления (рис. 9.1). Как оказалось, явление полного сцепления генов встречается в природе крайне редко. В настоящее время в природе только два вида имеют такой тип наследования: дрозофилы и тутовый шелкопряд, причем в обоих случаях это только гетерогаметный пол. В связи с этим становятся понятными результаты опытов Моргана: у гибридных самцов полное сцепление генов (отсутствие кроссинговера), поэтому образуется только 2 типа гамет, в результате оплодотворения образуются два фенотипических класса, без изменений повторяющих признаки родителей, В случае анализирующего скрещивания, где самка гибридна, группы сцепления самки могут нарушать-' ся явлением кроссинговера. При этом в гаметогенезе будут получаться гаметы, у которых отсутствовал кроссинговер (некроссоверы), й гаметы, которые образовались в результате кроссинговера (кроссоверы). После оплодотворения первые полностью повторили фенотипы родителей, а вторые образовали особи с новыми сочетаниями признаков. 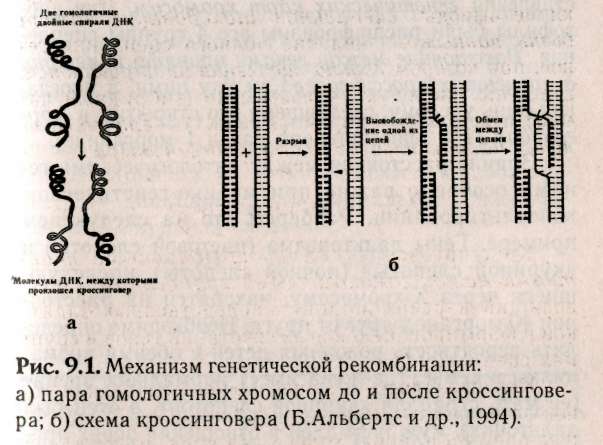 Кроссинговер бывает одинарный и множественный (в группе сцепления происходит обмен гомологичными локусами на одном или нескольких участках хромосом, что приводит к новым сочетаниям аллелей). Оказалось, что при неполном сцеплении генов процент кроссоверов (рекомбинант) кобщему числу потомков от анализирующего скрещивания — величина постоянная. На основании этого Морган предполагает, что процент кроссинговера отражает расстояние между генами: чем больше его величина, тем больше расстояние между генами (силы сцепления невелики), и, наоборот — чем меньше величина кроссинговера, тем меньше расстояние между генами (силы сцепления между генами сильнее). Возможность оценки расстояния между генами с помощью явления кроссинговера позволила Моргану и его сотрудникам обосновать методику составления генетических карт хромосом. Для дрозофилы были расшифрованы все 4 группы сцепления. Расстояние между генами принято измерять в процентах кроссинговера между ними, а иногда, как дань ученому, сделавшему это открытие, в морганидах (1% кроссинговера равен 1 морганиде). Знание расстояния между патологическими генами особенно важно при медико-генетическом консультировании. Разберем это на следующем примере. Гены дальтонизма (цветовой слепоты) и «куриной слепоты» (ночной слепоты), наследующиеся через Х-хромосому, находятся на расстоянии 40 морганид друг от друга. Необходимо определить вероятность рождения детей с обеими аномалиями в семье, где жена имеет нормальное зрение (ее мать страдала «куриной слепотой», а отец был дальтоник), муж нормален в отношении обоих признаков. Следует помнить, что эти патологии обуславливаются рецессивными аллелями. Для осуществления прогнозирования разберем, каковы генотипы родителей. У жены зрение нормально, но каждая из двух Х-хромосом несет по одной патологии (Х-хромосома, привнесенная яйцеклеткой матери, несет рецессивный аллель «куриной слепоты», а X-хромосома, привнесенная сперматозоидом отца, несет рецессивный аллель дальтонизма). Так как муж нормален по двум признакам, то его единственная Х-хромосома несет эти гены в доминантном состоянии. У него будет всего два типа гамет: Х-сперматозоиды будут нести здоровые аллели двух генов и У-сперматозоиды. У его жены будут образовываться четыре типа гамет (так как пара гомологичных хромосом будет вступать в крос-синговер, образуя рекомбинантные гаметы с частотой 40%). Общая вероятность образования гамет у жены равна 1, тогда вероятность образования пары рекомбинантных гамет равна 0,4, а некроссоверов (кроссинговер отсутствовал) — 0,6. В итоге, перемножая вероятности встречающихся при оплодотворении гамет, мы определяем вероятности рождения детей с разными фенотипами. Обозначим ген дальтонизма: D — нормальное цветовое зрение, d — дальтонизм, S — нормальное ночное зрение, s — «куриная слепота». р ♀Ds x ♂Ds dS здорова здоров
Все дочери в этом браке будут здоровы, но часть из них будет носительницей одной или двух патологий. 10% всех, детей (только мальчики) будут страдать двумя видами патологий, т.е. вероятность рождения таких детей 0,1. 30% детей (только мальчики) будут страдать какой-то одной из патологий поровну). Вероятность рождения здоровых-мальчиков равна 0,1 (или 10%). 9.3. Генетические карты Впервые Морган и его сотрудники использовали явление кроссинговера для составления генетических карт хромосом с использованием многочисленных мутантных генов. Генетическая карта хромосом — это схема взаимного расположения генов, находящихся в одной группе сцепления. Расстояние между генами на генетической карте хромосом определяют по частоте кроссинговера между ними (рис. 9.2). Единицей расстояния принято считать одну морганиду (один процент кроссинговера). Поскольку у человека способ составления генетических карт с помощью гибридологического анализа затруднен, разработаны другие способы определения локализации гена. Среди них: F-анализ расщепления и сцепления признаков в семьях (по родословным); так были картированы, например, локусы группы крови AB0; AAS — сравнение аминокислотных последовательностей белков; СН — изменение морфологии хромосомного участка в сочетании с характерным фенотипом; к этой группе относят анализ «ломких» (фрагильных) Достоверно идентифицировано около полутора тысяч генов человека, что составляет 1-5% от общего числа (рис. 9.3). У большинства из них обнаружены альтернативные аллельные формы. Около тысячи известных генов имеют хотя бы один из альтернативных аллелей, соответствующих какой-либо аномалии или заболеванию. Остальные гены кодируют белки группы крови, антигены, иммуноглобулины, сайтов в хромосомах. Фрагильные сайты полезны как маркеры-в семейных исследованиях сцепления; RE — рестрикционный анализ и реконструированные карты района; ОТ— центромерное картирование и др.  Достоверно идентифицировано около полутора тысяч генов человека, что составляет 1-5% от общего числа (рис. 9.3). У большинства из них обнаружены альтернативные аллельные формы. Около тысячи известных генов имеют хотя бы один из альтернативных аллелей, соответствующих какой-либо аномалии или заболеванию. Остальные гены кодируют белки группы крови, антигены, иммуноглобулины, ферменты и т.д. В настоящее время не существует принципиальных технических препятствий для получения полной карты генома человека, т.к. разработаны генно-инженерные подходы. Однако вследствие огромного размера изучаемой ДНК (см. раздел 4.1) на это уйдут многие годы. 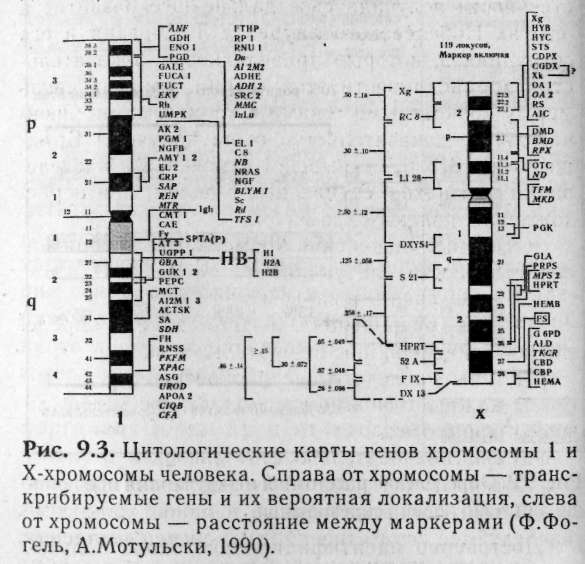 9.4. Хромосомная теория наследственности В 1902—1903 гг. два исследователя — Саттон (из США) и Бовери (из Германии) — независимо друг от друга предположили, что задатки (гены) расположены в хромосомах. Эта идея положила начало созданию хромосомной теории наследственности. Термин «ген» был предложен позже (в 1909 г. Иоганнсеном). Параллелизм в поведении генов и хромосом в процессе образования гамет убедительно говорил о том, что гены расположены в хромосомах. В 1910 г. хромосомная теория наследственности получила свое дальнейшее развитие в опытах Нобелевского лауреата Т.Моргана и его сотрудников, которые привели новые доказательства справедливости этой теории, показав связь между конкретными генами и хромосомами. Окончательные доказательства были получены Брид-жесом в 1913 г., открывшем изменения в наследовании признаков, сцепленных с полом, при нерасхождении половых хромосом. Основные положения хромосомной теории наследственности следующие: гены локализованы в хромосомах; гены расположены в хромосоме в определенной линейной последовательности; гены наследственно дискретны; каждый ген имеет определенное место (локус) в хромосоме; гены относительно стабильны; гены могут изменяться (мутировать); гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов; признаки, зависящие от сцепления генов, наследуются совместно; сцепление генов может нарушаться процессом кроссинговера, в результате образуются рекомбинантные хромосомы; частота кроссинговера является функцией расстояния, между генами: чем больше расстоя-, ние, тем больше величина кроссинговера (прямая зависимость); частота кроссинговера зависитют силы сцеп-, ления между генами: чем сильнее сцеплены.гены, тем меньше величина кроссинговера (обратная зависимость); сцепление генов и кроссинговер позволяют производить картирование хромосом. На современном этапе развития хромосомная теория наследственности получила дальнейшее развитие благодаря изучению тонкой структуры и функции генов про- и эукариот. Эволюционировали наши представления о гене как о неделимой структуре. Открыты дробные структуры эукариоти-ческих генов, механизмы их регуляции и т.д. Созданы искусственные гены. Все эти знания позволили сформулировать один из фундаментальных принципов генетики о единстве дискретности и непрерывности генетического материала. 9.5. Задание 1. У человека катаракта и многопалость вызываются доминантными аллелями двух сцепленных генов. Здоровая женщина выходит замуж за мужчину, страдающего этими признаками (многопалость унаследовал от отца, а катаракту от матери). Оцените вероятность того, что их ребенок будет: а) одновременно страдать обеими аномалиями; б) страдать одной из двух аномалий; в) полностью здоров? Допустите, что кроссинговер не происходит. 2. У человека резус-Положйтёльность и элипто-цитоз Определяются сцепленными доминантными! генами (расстояние 3 мортаниды). В брак вступает мужчина с элиптоцитозом и резус-положительным фактором (мать которого имела отрицательный резус-фактор, а отец был болен элиптоцитозом) и здоровая женщина, имеющая отрицательный резус-фактор. Определите вероятность возможных фенотипов детей в этой семье. 3. Гены А и В относятся к одной группе сцепления, расстояние между генами 40 морганид. Оцените вероятность рождения детей в браке, где оба родителя дигетерозиготны, при этом женщина получила доминантные гены от отца, а мужчина — один — от отца, другой — от матери. 4. В брак вступают родители, страдающие катарактой и полидактилией (оба дигетерозиготны). Оба родителя получили катаракту от матерей, а полидактилию — от отцов. Каков прогноз в отношении здоровья детей, если допустить, что кроссинговер не наблюдается? 5. Гены цветовой и ночной слепоты рецессивны, сцеплены и находятся в Х-хромосоме на расстоянии 50 морганид. Девушка с нормальным зрением (отец которой страдал двумя формами слепоты) выходит замуж. Определите вероятность появления возможных фенотипов у детей в этой семье, если: а) юноша здоров? б) страдает двумя формами слепоты? 6. У здоровой женщины родились сын-гемофилик с нормальным зрением и дальтоник-дочь с нормальной свертываемостью крови. Определите возможные генотипы родителей указанных детей. Оцените вероятность рождения следующего ребенка здоровым. 7. Классическая гемофилия и дальтонизм наследуются рецессивно, сцепленно с Х-хромосомой. Расстояние между генами 9,8%. Женщина, мать которой была дальтоник, а отец гемофилик, вступила в брак со здоровым мужчиной. Определите I вероятность рождения в этой семье детей с двумя аномалиями одновременно. 8. У человека мышечная дистрофия Дюшена и полная цветовая слепота (протанопия) наследуются сцепленно с полом, рецессивно. Здоровая женщина с нормальным зрением (отец страдал мышечной дистрофией, а мать протанопией) вышла замуж за мужчину, страдающего обоими заболеваниями. Какой прогноз рождения здоровых детей в этом браке? Примечание. При решении задач № 5—8 необходимо помнить, что кроссинговер в половых хромосомах у мужчин практически не наблюдается. |