Глик Молекулярная биотехнология. Глик Б., Пастернак Дж. Молекулярная биотехнология. Принципы и применение. Пер с англ. М. Мир, 2002. 589 с
 Скачать 9.74 Mb. Скачать 9.74 Mb.
|

| |||||||||||||||||
| Рис. 16.4. Разные типы биореакторов (упрощенная схема), А. Реактор с механическим перемешиванием . Б. Барботажная колонна. В. Эрлифтный реактор с внутренней рециркуляцией. Г. Эрлифтный реактор с внешней системой рециркуляции. Стрелки -- направление потока культуральной среды. | 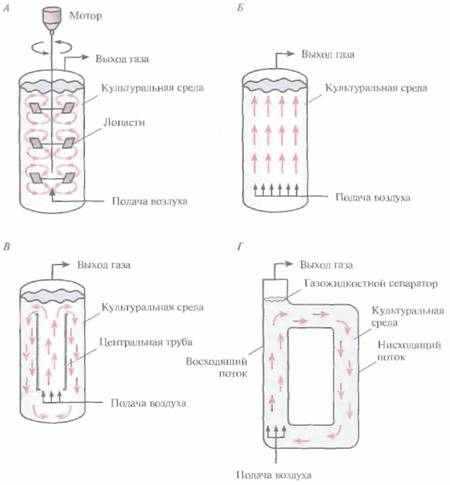 |
358 ГЛАВА 16
нерном языке, обладают высоким объемным коэффициентом массообмена, kLa) • уже давно используются для выращивания различных микроорганизмов.
В реакторах с механическим перемешиванием газ (как правило, воздух) подают в культуральную среду под давлением через разбрызгиватель - - кольцо с множеством маленьких отверстий либо трубку с одним отверстием. В первом случае образуются мелкие пузырьки воздуха и обеспечивается их более равномерное распределение, однако разбрызгиватели в виде трубок используются чаще, поскольку они реже закупориваются. Для равномерного распределения газа по всему объему биореактора используются мешалки — одна или несколько. Они разбивают крупные пузырьки воздуха, разносят их по всему реактору и увеличивают время пребывания в культуральной среде. При сильном перемешивании средний размер пузырьков в больших биореакторах практически не зависит от размера отверстий в разбрызгивателе. Эффективность распределения газа зависит прежде всего от типа мешалки, числа оборотов и физико-химических свойств среды. Если размер биореактора слишком велик, а газ, поступающий из разбрызгивателя, распределяется по объему неравномерно, то даже при энергичном перемешивании гомогенизировать среду не удается.
Многие культуральные среды весьма агрессивны, и во избежание коррозийного или механического повреждения стенок биореактора его обычно изготавливают из нержавеющей стали или стекла. Стеклянные части чаще используют только в лабораторных биореакторах емкостью меньше 50 л.
Размер биореактора лимитируется его способностью эффективно отдавать тепло, выделяемое микроорганизмами в ходе метаболизма и высвобождаемое в результате перемешивания. Если теплоотдача недостаточна, температура среды может превысить критическую, что уменьшит выход продукта. Для отвода тепла используют охлаждающую рубашку или змеевики, помещаемые внутрь реактора. Внутреннее охлаждение более эффективно, однако змеевики часто покрываются слоем растущих клеток, что затрудняет охлаждение, а иногда мешает интенсивному перемешиванию культуральной среды.
Большую опасность представляет загрязнение ферментера грибами или бактериями. Поэтому биореакторы конструируют таким образом, чтобы их можно было стерилизовать; обычно для этого используют пар под давлением. Внутри реактора не должно быть «мертвых зон», недоступных для пара во время стерилизации. Обработке подлежат все клапаны, датчики, входные и выходные отверстия. При конструировании перед инженерами зачастую возникает проблема: использовать максимальное число датчиков для полного контроля за процессом ферментации или ограничиться их минимальным набором, чтобы легче было поддерживать стерильность.
При интенсивном перемешивании культуральной среды в процессе ферментации часто происходит ее вспенивание. Это может привести к переувлажнению фильтра в отверстии, через которое воздух выходит из биореактора, и уменьшению его потока, а также к попаданию в реактор посторонних микроорганизмов. Для контроля пенообразования используют химические пеногасители или механические сбиватели пены. Однако в присутствии химических реагентов может ухудшаться перенос кислорода, а иногда происходить ингибирование клеточных ферментов, что уменьшает скорость роста микроорганизмов. Кроме того, если пеногасители не удалять, они могут загрязнять конечный продукт. Проблему вспенивания можно решить, если оставить в верхней части биореактора достаточно большое пустое пространство, в котором лопались бы пузырьки воздуха. Правда, в этом случае рабочий объем реактора уменьшится примерно на 25%,
Все эти соображения относятся и к «пневматическим» реакторам типа барботажньгх колонн и эрлифтных биореакторов. Таким образом, обеспечение стерильности, постоянства pH и температуры — ключевые требования при любом способе культивирования независимо от конструкции биореактора.
Конструкционные особенности барботажных колонн и эрлифтных биореакторов дают им некоторые преимущества перед реакторами с механическим перемешиванием, «Пневматические» реакторы более экономичны, поскольку
Промышленный синтез белков при участии рекомбинантных микроорганизмов 359
перемешивание в них происходит с помощью восходящего потока воздуха (или другого газа в случае анаэробных микроорганизмов), а не механической мешалки, потребляющей много энергии. Кроме того, в отсутствие механической мешалки исключается и один из путей проникновения в биореактор посторонних микроорганизмов. В пневматических биореакторах в культуральной среде не возникает столь сильных гидродинамических возмущений (сдвига слоев жидкости друг относительно друга), при этом в эрлифтных биореакторах перемешивание происходит более равномерно по всему объему. Уменьшение сдвиговых эффектов очень важно по следующим причинам:
* клетки рекомбинантных микроорганизмов более хрупки, чем нетрансформированные клетки, поскольку часть их энергетических ресурсов расходуется на синтез чужеродных белков и в результате образуется менее прочная клеточная стенка
* самый распространенный ответ клетки на внешнее воздействие — уменьшение количества всех синтезируемых белков, в том числе и рекомбинантных
* под действием сдвиговых эффектов могут изменяться физические и химические свойства клеток, что затруднит дальнейшую работу с ними. Например, может увеличиться количество полисахаридов на поверхности клеток, что приведет к ухудшению условий их выделения и лизиса, а также затруднит очистку рекомбинантного белка.
В барботажных колоннах воздух подается под высоким давлением в нижнюю часть биореактора; по мере подъема маленькие пузырьки воздуха объединяются, что приводит к неравномерному его распределению. Кроме того, подача воздуха под высоким давлением может привести к слишком сильному пенообразованию. Все это ограничивает универсальность данных конструкций и сужает диапазон реализуемых технологических условий, а также уменьшает возможный размер барботажных колонн.
Эрлифтные биореакторы могут использоваться как в экспериментальных установках, так и в целях промышленной ферментации. Газ в
них подается в нижнюю часть вертикального канала. Поднимаясь, он увлекает за собой жидкость к верхней части канала — газожидкостному сепаратору, и здесь частично выходит в воздух. Более плотная деаэрированная жидкость опускается по другому вертикальному каналу ко дну реактора, и процесс повторяется. Таким образом, культуральная среда вместе с клетками непрерывно циркулирует в биореакторе.
Эрлифтные биореакторы бывают двух основных типов. В первом случае реактор представляет собой одну емкость с центральной трубкой, которая обеспечивает циркуляцию жидкости (реакторы с внутренней рециркуляцией) (рис. 16.4, В). Во втором культуральная среда проходит через отдельные, независимые каналы (реактор с внешней рециркуляцией) (рис. 16.4, Г). Конструкция эрлифтных реакторов с внутренней рециркуляцией проще, но если уж реактор построен, его объем и скорость циркуляции остаются неизменными. Напротив, биореактор с внешней рециркуляцией можно модифицировать и создавать разные условия ферментации.
Эрлифтные биореакторы, вообще говоря, более эффективны, чем барботажные колонны, особенно в случае суспензий микроорганизмов с большой плотностью или вязкостью. Перемешивание в них более эффективно и проблема слипания пузырьков не столь велика. В особенно больших эрлифтных ферментерах, таких как ферментер на ] 500 000 л фирмы IC1 (Англия), сконструированный для получения белков одноклеточных микроорганизмов, для прохождения клетками полного цикла в реакторе требуется весьма значительное время. Чтобы обеспечить их субстратами на все время их перемещения с током жидкости, субстраты вводились по всей длине реактора сразу во многих точках.
Типичные крупномасштабные системы ферментации
Рекомбинантные микроорганизмы широко используются для получения разнообразных белковых продуктов, применяющихся в медицине (например, инсулина), а также в качестве своего рода «фабрик» по производству имеющих коммерческую ценность метаболитов (например, антибиотиков). Белки синтезируются наиболее
360 ГЛАВА 16
интенсивно в период от середины экспоненциальной фазы до ее завершения, а метаболиты — в период замедления роста и в стационарной фазе. Все это должно учитываться при выборе параметров крупномасшабных процессов ферментации.
Оптимизация синтеза необходимого продукта — серьезная научная проблема. Если речь идет о белках, то для ее решения обычно используют клонированные гены, находящиеся под контролем сильных регулируемых промоторов. Вначале полагали, что для получения нужного количества продукта будет достаточно конститутивной экспрессии клонированного гена. Однако опыт показал, что при непрерывной транскрипции и трансляции клонированного гена истощаются все энергетические ресурсы клетки и ее рост замедляется. Чтобы приурочить экспрессию клонированного гена к определенной фазе роста, можно использовать механизм индукции. Для этого вначале выращивают клетки в оптимальных условиях до относительно высокой плотности, а затем индуцируют транскрипцию, либо изменяя температуру, либо добавляя в среду тот или иной химический индуктор в зависимости от природы промотора (например, изопропил-ß-тиогалактопиранозид ).
Двухступенчатая ферментация в большом биореакторе (> 100 л) встречается с определенными трудностями, поскольку технически очень сложно быстро повысить температуру (обычно с 30 до 42 °С) в большом объеме или обеспечить быстрое и равномерное распределение химического индуктора. Эту проблему можно решить, если использовать два сообщающихся биореактора (двухступенчатая ферментация): клетки выращивают в одном из них, а индукцию осуществляют в другом. Это позволяет оптимизировать процессы роста и индукции по отдельности и увеличить количество продукта, синтезируемого за единицу времени.
Двухступенчатая ферментация в тандемных эрлифтных биореакторах
Штамм E. соli NM989, несущий ген ДНК-лигазы Т4 под транскрипционным контролем промотора pLи температурочувствительного репрессора cI, выращивали и индуцировали в двухступенчатом эрлифтном биореакторе (рис. 16.5). Ген
ДНК-лигазы был встроен в хромосомную ДНК, что снимало все проблемы, связанные с нестабильностью плазмид в ходе длительной ферментации. Клетки выращивали при 30 °С в эрлифтном биореакторе с внешней рециркуляцией и рабочим объемом 10 л. В этих условиях ген ДНК-лигазы не экспрессировался. Для индукции при 42 °С использовали эрлифтный биореактор с внешней рециркуляцией и рабочим объемом около 5 л. Биореакторы были соединены трубкой с насосом, который обеспечивал непрерывность подачи суспензии из первого биореактора во второй. Клеточную суспензию, достигшую определенной плотности, удаляли из биореактора, где проходила индукция, и подвергали дальнейшей обработке.
Максимальная удельная скорость роста культуры (µmax) составляла примерно 0,66 ч1 в первом биореакторе и 0,54 ч1 во втором, что соответствовало времени удвоения 63 и 77 мин. Свежую среду непрерывно добавляли в ферментер, где росли клетки, со скоростью 2 л/ч, а из ферментера, где происходила индукция, отбирали такой же объем суспензии. Поскольку рабочие объемы биореакторов различались, клетки находились примерно 5 ч в биореакторе, где происходил рост, и 2 ч в биореакторе, где осуществлялась индукция. Различие во времени пребывания клеток в биореакторах было необходимо для оптимизации числа клеток, выхода продукции и стабильности ДНК-лигазы. Само время пребывания клеток в разных реакторах можно варьировать изменением их относительного рабочего объема и объема поступающих в первый биореактор питательных веществ.
Использованный в этой работе эрлифтный ферментер с двойной наружной рециркуляцией (рис. 16.5) позволил упростить регуляцию относительных рабочих объемов ферментеров, а также повысить гибкость системы (обеспечивать разные условия роста для разных популяций рекомбинантных клеток). При синтезе ДНК-лигазы наилучшие результаты были получены при ежеминутном поступлении примерно 33 мл клеточной суспензии из первого биореактора во второй. Это эквивалентно всего 0,67% объема биореактора, где осуществлялась индукция, что обеспечивало практически мгновенный подъем температуры всей поступающей суспензии с 30
Промышленным синтез белков при участии рекомбинантных микроорганизмов 361
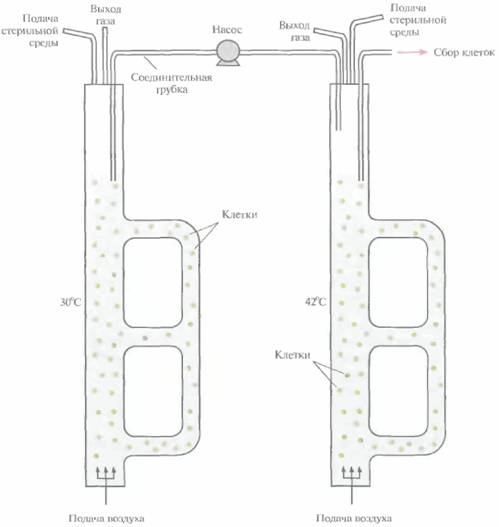 |
| Рис. 16,5. Система из двух эрлифтных биореакторов, использующаяся для температурной индукции синтеза белкового продукта. Из ферментера, в котором осуществляется культивирование при 30 °С (слева), клетки поступают в ферментер с температурой 42 °С (справа}, где происхоит индукция. У обоих биореакторов имеется двойное внешнее рециркуляционное устройство, оснащенное задвижками. Изменяя положение задвижек, можно создавать рабочий объем, оптимальный для данных условии. |
до 42 °С. Для поддержания роста клеток, находящихся во втором биореакторе, в экспоненциальной фазе в него непрерывно добавлялось нужное количество питательных веществ в концентрированной форме. Это предотвращало расщепление ДНК-лигазы протеолитическими ферментами, которые обычно синтезируются клетками в фазе замедления и в стационарной фазе.
При росте в таком двухступенчатом биореакторе непрерывного действия культура штамма E. coliNM989 может достигать плотности 4 г (сухого вещества) на 1 л, а на долю ДНК-лигазы Т4 может приходиться до 4% суммарного белка в клетке, что соответствует примерно 25 000 ЕД ферментативной активности на 1 г (сухого вещества). Вообще же с помощью описанного подхода можно синтезировать примерно 100 000 ЕД
362 ГЛАВА 16
ферментативной активности на 1 л культуры, т. е. до 4 800 000 ЕД в сутки. С учетом того что после очистки фермента уровень активности уменьшается до 20% и что стоимость единицы активности равна примерно 0,25 долларов, получаем, что ежедневно можно синтезировать количество фермента на сумму 240 000 долларов. Мы не учли всех затрат на само производство белка, однако ясно и так, что прибыль от реализации ценных продуктов, полученных при непрерывной ферментации в биореакторах малого или среднего размера, значительно превышает затраты.
Двухступенчатая ферментация в одном реакторе с механическим перемешиванием
Трехкомпонентный рекомбинантный белок AGßgal, использующийся при проведении иммунологических тестов, синтезировали в промышленных масштабах в одном биореакторе с механическим перемешиванием. Ген этого белка был сконструирован методами генной инженерии и содержал сегменты, кодирующие пять сайтов связывания иммуноглобулина G (JgG) А-белка Staphylacoccus aureus, два сайта связывания IgG G-белка штамма G148 Streptococcusи ß-галактозидазу Е. coli. Он находился под контролем промотора pR бактериофага λ, регуляция которого осуществляется так же, как регуляция pL-промотора, и был встроен в плазмиду, несущую ген устойчивости к ампициллину; этой конструкцией трансформировали клетки Е. соli. Штамм с плазмидой, несущей ДНК AGßgai, содержал вторую плазмиду, несущую ген температурочувствительного белка-репрессора сIи ген устойчивости к канамицину.
Культуру в объеме 5 л выращивали при 30 °С в присутствии ампициллина и канамицина, с тем чтобы обеспечить условия для сохранения обеих плазмид, а затем использовали в качестве посевного материала для инициации роста культуры без антибиотиков при 30 °С в реакторе с рабочим объемом 45 л. Суспензия клеток из 45-литрового ферментера в свою очередь служила посевным материалом для культивирования в биореакторе на 600 л, в котором клетки продолжали выращивать при 30 'С без антибиотиков (рис. 16.6). (Вообще говоря, для уменьшения стоимости процесса антибиотики при крупно-
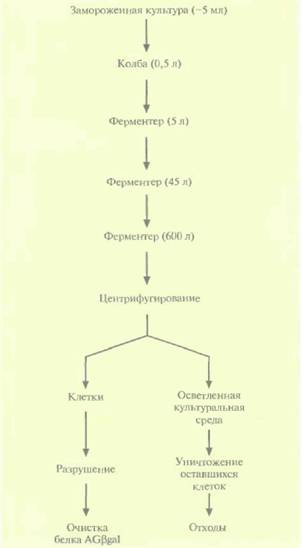 |
| Рис. 16.6. Схематическое представление промышленного синтеза белка AGßgai. В скобках указан объем культуральной среды на каждом этапе. На ее долю приходится от 60 до 75% объема соответствующих биореакторов (ферментеров). |
масштабном культивировании в среду не добавляют.) Как только плотность культуры в биореакторе на 600 л достигала примерно 4 г/л, температуру повышали с 30 до 40 °С, чтобы индуцировать экспрессию гена белка AGßgal. На повышение температуры в этих условиях уходило около часа. Индукцию проводили при 40, а не при 42 °С, по-
Промышленный синтез белков при участии рекомбинантных микроорганизмов 363
скольку при более низкой температуре синтезировалось такое же количество белка AGßgal, но клетки могли расти в течение более длительного периода. Иными словами, при более низкой температуре индукции {40 °С) выход белкового продукта был больше.
Удельная активность белка AGßgal в течение 2 ч после начала индукции повышалась, а затем снижалась. Возможно, это было связано с синтезом протеаз клетками, перешедшими в фазу замедленного роста или в стационарную фазу. Кроме того, примерно 50% клеток, росших в течение 4 ч при 40 °С, утратили плазмиду. Но даже при этом после 4-часового культивирования при 40 °С на долю AGßgal-белка приходилось примерно 20% всей массы сухого вещества. Учитывая все это, можно не интегрировать гены белка AGßgal и репрессора cl в хромосомную ДНК хозяйской клетки Е, coliс целью увеличения выхода продуктов.
Периодическая ферментация и периодическая ферментация с добавлением субстрата
В некоторых случаях для достижения высокой плотности культуры и получения больших количеств продукта достаточно проводить ферментацию в обычном периодическом режиме. В одном из экспериментов плазмиду, несущую ген гибридного белка, одним из компонентов которого был пептид инсулина В, помещали под контроль trp-промотора E, соliи вводили в trp-штамм Е. соli; трансформированные клетки
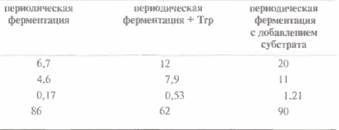
культивировали в среде с разным содержанием триптофана. При высоких концентрациях последнего гибридный белок не синтезировался; последующее поглощение триптофана из среды растущими клетками приводило к индукции синтеза необходимого белка. Добавление триптофана в среду приводило к увеличению количества как биомассы, так и синтезируемого белка, а использование периодических ферментеров с добавлением постоянного количества субстрата еще более усиливало этот эффект (табл. 16.1).
Сбор клеток
Чтобы очистить продукт ферментации, нужно прежде всего отделить клетки от культуральной среды. Сбор генетически модифицированных и исходных, нетрансформированных клеток можно проводить одними и теми же методами. Однако трансформированные клетки часто обладают другими физиологическими свойствами (они имеют другой размер или синтезируют внеклеточные полисахариды), и в результате условия, оптимальные для сбора нетрансформированных клеток, могут не подходить для клеток, синтезирующих чужеродный белок.
Для выделения клеток из больших объемов культуральной среды часто используют высокоскоростное центрифугирование. Для этого сконструированы специальные высокоскоростные центрифуги полунепрерывного действия. Суспензию клеток непрерывно подают в барабан работающей центрифуги, клетки концент-
| Таблица 16. 1. Синтез рекомбинантного белка, одним из компонентов которого является пептид инсулина В, при периодической ферментации и при периодической ферментации с добавлением субстрата1)2) | |
| Конечный показатель | Выход |
| |  |
 | |
| 1) Поданным работы Cosset et al,, AppL MikrobioL Biotechnol. 39; 541-546, 1993. 2) Количество биомассы измеряется в граммах сухого вещества на литр культуры. Процедура «Периодическая ферментация + Тrр» означает, что к культуре добавлено 0,1 г триптофана. При периодической ферментации с добавлением субстрата в среду добавляли 0,1 г триптофана каждые 2 ч — в обшей сложности пять раз в течение 10 ч. При добавлении большего количества триптофана не увеличивались ни биомасса, ни количество синтезированного белка. | |
364 ГЛАВА 16
рируются в нем, а осветленная среда удаляется, Когда барабан заполняется осажденными клетками, центрифугу останавливают и клетки собирают. Основное неудобство данного способа -необходимость останавливать процесс, а затем снова начинать его. Кроме того, недостатками являются высокая стоимость оборудования и потребляемой им энергии, вероятность утечки микроорганизмов в окружающую среду, невозможность полного удаления клеток из среды.
Альтернативный метод выделения клеток из культуральной среды — фильтрация через мембрану. К сожалению, при обычной фильтрации клетки со временем забивают поры мембранного фильтра, накапливаются на его поверхности, и в результате скорость процесса быстро снижается (рис. 16.7, А). Фильтрацию можно ускорить, проводя ее под давлением, но это лишь временный эффект; клетки все равно будут накапливаться на поверхности мембраны, а кроме
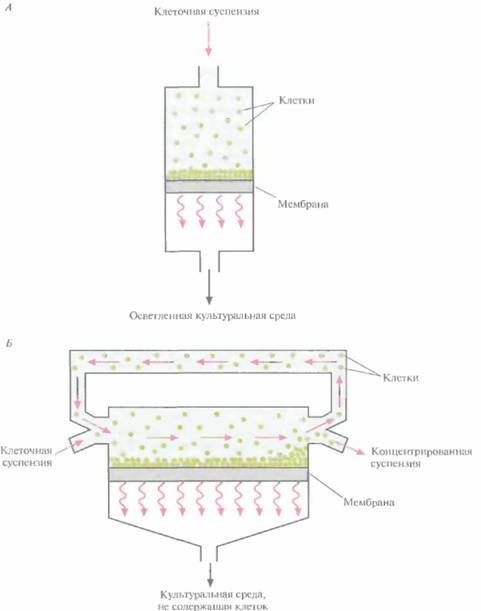 | Рис. 16.7. Способы фильтрации, применяющиеся для сбора клеток. A. Фильтрация с необратимым забиванием фильтра. Б. Фильтрация с параллельным потоком клеточной суспензии. Стрелки — направление потока. |
Промышленный синтез белков при участии рекомбинантных микроорганизмов 365
того, под давлением они образуют более плотный и менее проницаемый слой.
Чтобы решить эту проблему, клеточную суспензию пропускают с высокой скоростью параллельно поверхности мембраны (рис. 16.7, Б), так что через мембрану за один раунд проходит только небольшая часть циркулирующей жидкости. Остальная ее часть очищает мембрану от накопившихся клеток (см. рисунок), и в результате скорость фильтрации падает не так быстро, как при необратимом забивании фильтра. После многочисленных раундов фильтрации через мембрану проходит почти вся культуральная среда. Этот метод используется пока только в лаборатории; в промышленных процессах для сбора клеток применяют центрифугирование.
Дальнейшие действия зависят от природы и локализации продукта. Если продукт представляет собой белок, находящийся в культуральной среде, то среду концентрируют, а белок очищают хромотографическими или другими методами. Если продукт — это низкомолекулярное соединение, находящееся в культуральной среде, то используют соответствующие методы экстракции. Наконец, если продукт имеет внутриклеточную локализацию, то прежде чем очищать его, клетки разрушают.
Разрушение клеток
Для разрушения клеток используют разнообразные химические, биологические и физические методы. Все процедуры должны быть достаточно жесткими, чтобы разрушить клеточную стенку, и в тоже время достаточно мягкими, чтобы исключить денатурацию белка. А поскольку клеточные стенки у разных микроорганизмов состоят из разных полимеров, никакого универсального метода их разрушения не существует.
• У грамположительных бактерий клеточная стенка состоит из толстого пептидогликано-вого слоя N-ацетил глюкозам и на и остатков N-ацетилмурамовой кислоты, соединенных пептидными мостиками.
• У грамотрицательных бактерий клеточная стенка тоньше и покрыта снаружи слоем липидов.
• Стенка дрожжевых клеток состоит из плотного слоя частично фосфорилированных маннанов и ß-глюканов.
* Низшие грибы имеют многослойные клеточные стенки, состоящие из α- и ß-глюканов, гликопротеидов и хитина.
Состав и прочность клеточной стенки зависят от условий культивирования, скорости роста клеток, фазы, на которой они собираются, условий хранения сконцентрированных клеток и от того, экспрессировал ли выделенный микроорганизм клонированный ген.
Химические методы разрушения клеточных стенок включают обработку щелочью, органическими растворителями или детергентами. Если белковый продукт не разрушается при pH от 10,5 до 12,5, то можно без труда и дешево лизировать большие количества бактериальных клеток. Например, рекомбинантный гормон роста человека очень просто выделить из клеток Е. сой обработкой гидроксидом натрия при pH 11. После обработки щелочью не остается практически ни одной жизнеспособной клетки, что автоматически решает проблему утечки рекомбинантных микроорганизмов. Обработка органическими растворителями — это простой и недорогой способ разрушения клеток, который используется для выделения ферментов из дрожжей. Однако, чтобы убедиться в том, что в подобранных условиях, белковый продукт не денатурирует, необходимо провести предварительное тестирование. Под действием детергентов в мембранах бактериальных клеток образуются поры, через которые белки и другие молекулы выходят из клетки. К сожалению, детергенты дороги, в большинстве случаев в их присутствии белки денатурируют, а кроме того, они могут загрязнять конечный продукт.
Основным биологическим методом разрушения клеток микроорганизмов является лизис с помощью ферментов. Так, лизоиим яичного белка легко гидролизует клеточные стенки грамположительных бактерий. Для разрушения клеточных стенок грамотрицательных бактерий используют лизоцим и этилендиаминтетрауксусную кислоту (ЭДТА), а клеточные стенки дрожжей гидролизуют с помощью одного или
366 ГЛАВЛ 16
нескольких ферментов: β-1,3-глюканазы, β-1,6-глюканазы, манназы и хитиназы. Ферментативная обработка высокоспецифична, а лизис проходит в мягких условиях. Пока использование ферментов для лизиса клеток сдерживается их высокой стоимостью, но с применением рекомбинантных микроорганизмов для промышленного синтеза ферментов, разрушающих клеточные стенки, эта проблема будет решена.
Клетки можно разрушить и физическими методами: немеханическими (например, с помощью осмотического шока или быстрого многократного замораживания и оттаивания) или механическими (обработкой ультразвуком, с помощью шаровой мельницы, гомогенизации под давлением, соударения). Обычно после обработки немеханическими методами многие клетки остаются неповрежденными. Напротив, механическое разрушение высокоэффективно, что делает его более привлекательным. Особенно часто ультразвуковые излучатели, генерирующие высокочастотные звуковые волны, используют для обработки малых объемов. Клетки разрушаются при этом под действием гидродинамических сил (сдвига слоев жидкости друг относительно друга, кавитации и т. д.).
Для разрушения большого количества клеток обычно используют шаровые мельницы. Концентрированную клеточную суспензию заливают в камеру высокоскоростной шаровой мельницы, заполненную инертным абразивным материалом (например, стеклянными шариками диаметром <1 мм). Содержимое быстро перемешивают с помощью лопастей, насаженных на ось. Большинство клеток разрушается под действием сдвиговых напряжений, возникающих в результате быстрого движения шариков. Условия оптимального разрушения клеток можно подобрать, варьируя число и форму лопастей, скорость перемешивания, размер шариков, их число, концентрацию клеток, геометрию камеры и температуру. Приборы такого типа успешно использовались для разрушения клеток самых разных микроорганизмов. С их помощью можно легко разрушать клетки как нерекомбинантных, так и рекомбинантных микроорганизмов.
При гомогенизации под высоким давлением концентрированную клеточную суспензию продавливают через небольшое отверстие под высоким давлением, а затем давление резко сбрасывают, что и вызывает лизис. Условия обработки можно оптимизировать применительно к разным микроорганизмам. Для этого изменяют рабочее давление, размер и форму отверстия, температуру клеточной суспензии, число продавлива-ний.
Еще один механический метод разрушения клеток -- соударение. Клеточную суспензию большой вязкости направляют под давлением на неподвижную поверхность или навстречу потоку другой суспензии. В месте соприкосновения выделяется большое количество энергии, разрушающей клетки. Таким способом с помощью устройства под названием Microfluidizer за один прием была разрушена большая часть клеток Е. collв двух встречных потоках суспензии. Однако для разрушения клеток других микроорганизмов может понадобиться большее число раундов. В отличие от гомогенизаторов под высоким давлением и высокоскоростных шаровых мельниц, в которых, как правило, используются концентрированные клеточные суспензии, данное устройство пригодно для обработки любых суспензий. Как показали предварительные исследования, активность клеточных белков уменьшается при разрушении клеток по этой методике лишь незначительно. А если обработать суспензию клеток небольшим количеством лизоцима, а затем использовать устройство Microfluidizer в режиме пониженного по сравнению с обычным давления и при небольшой вязкости, то сохранится активность некоторых лабильных белков, инактивирующихся при высоком давлении.
Дальнейшая обработка
После разрушения клеток их осколки удаляют либо низкоскоростным центрифугированием больших объемов, либо микрофильтрацией через мембрану. Белковый продукт осаждают из грубого или осветленного лизата органическими растворителями (спиртом или ацетоном) или сульфатом аммония. Достигаемое при этом обогащение — 2—5 раз. К сожалению, дороговизна
Промышленный синтез белков при участии рекомбинантных микроорганизмов 367
агентов, использующихся для осаждения, может значительно увеличить стоимость процесса. В качестве альтернативы для концентрирования и выделения суммарных белков можно использовать ультрафильтрацию с параллельным потоком через мембраны с меньшим средним размером пор, чем у мембран, применяющихся для концентрации клеток или удаления их осколков (рис, 16.7, Б). Этот подход пока находится в стадии разработки, однако уже ясно, что он пригоден для работы с объемами от одного до нескольких тысяч литров, процесс может идти непрерывно (что позволяет уменьшить размеры установки) и обеспечивать 10—100-кратное обогащение (в зависимости от размеров и свойств выделяемого белка).
Необходимая степень очистки белкового продукта зависит от того, где его намереваются использовать. В одних случаях это может быть довольно грубый препарат, в других (например, если речь идет о белках, использующихся в медицине) — препарат высокой степени чистоты.
Некоторые белки, синтезирующиеся в клетках в избыточном количестве, образуют нерастворимые частицы (тельца включения). После разрушения клеток их легко можно отделить от большинства других клеточных компонентов. Вначале исследователям не удавалось дезагрегировать выделенные тельца включения так, чтобы при этом не произошла необратимая денатурация белков, но позже были разработаны методы, позволяющие ренатурировать рекомбинантный белок и восстановить его активность. Ясно, что все эти дополнительные процедуры увеличивают стоимость процесса очистки.
Солюбилизация белков
В некоторых случаях при гиперпродукции рекомбинантных белков образуются как растворимые, так и нерастворимые продукты, что усложняет процедуру очистки. Например, при экспрессии в клетках Е. coliгена инсулинопо-добного фактора роста I (IGF-I) человека мол. массой 7,6 кДа примерно 90% рекомбинантных молекул локализуется в периплазме, а 10% секретируется. Чтобы выделить растворимую и нерастворимую формы IGF-I из периплазмы, для солюбилизации нерастворимой формы in situ добавляли мочевину и дитиотрейтол до высоких концентраций при щелочном pH. При этом клетки погибали, но не разрушались, так что цитоплазматические белки оставались внутри клеток. В результате образовывался очень вязкий раствор, что затрудняло осаждение клеток и их осколков центрифугированием. Чтобы решить эту проблему, разработали процедуру двухфазной жидкостной экстракции, позволяющую разделять растворимые и нерастворимые продукты. И солюбилизация in situ, и двухфазная жидкостная экстракция высокоэффективны; с их помощью можно выделить от 80 до 95% IGF-I из культуры объемом от 10 до 1000 л.
ЗАКЛЮЧЕНИЕ
Для крупномасштабного культивирования рекомбинантных микроорганизмов в промышленных биореакторах (>1000 л) недостаточно просто экстраполировать условия роста в лабораторных ферментерах (0,1 — 1,0 л). При конструировании промышленных биореакторов необходимо учитывать такие параметры, как температура, pH, скорость и характер перемешивания, потребность аэробных организмов в кислороде, количество питательных веществ.
Ферментацию можно проводить по-разному. При периодической ферментации посевной материал вводят в свежую культуральную среду и проводят культивирование, не добавляя субстрат до тех пор, пока количество нужного продукта не достигнет максимума. В этих условиях рост культуры проходит шесть этапов; латентную фазу, фазу ускорения, логарифмическую (log) фазу, фазу замедления, стационарную фазу и фазу отмирания. Больше всего белков синтезируется во время логарифмической фазы, а многие низкомолекулярные продукты — во время стационарной. При таком способе ферментации необходимо тщательно следить за тем, чтобы клетки были собраны в нужное время. При периодической ферментации с добавлением субстрата в биореактор добавляют свежую культуральную среду через разные интервалы времени, как правило для того, чтобы продлить логарифмическую фазу. Непрерывная ферментация предполагает добавление свежей среды в течение всего процесса и одновременное удаление клеток и отработанной среды.
368 ГЛАВА 16
Каждая из этих систем ферментации имеет свои недостатки и достоинства, которые нужно учитывать, применяя ее для промышленного синтеза рекомбинантных продуктов. Несмотря на то что непрерывная ферментация применяется в промышленном масштабе не очень широко, этот способ имеет ряд преимуществ и в будущем, по-видимому, получит большее распространение.
Один из подходов к увеличению количества рекомбинантного белкового продукта состоит в максимальном увеличении плотности культуры трансформированных клеток, синтезирующих данный продукт. Для достижения этой цели лучше всего использовать режим периодической ферментации с добавлением субстрата,
Все биореакторы можно отнести к одному из трех основных типов: реакторы с механическим перемешиванием, барботажные колонны, эрлифтные реакторы. В настоящее время в промышленности чаше всего используются биореакторы первого типа, но появляется интерес и к эрлифтным биореакторам. Механическое перемешивание обеспечивается с помощью механической мешалки, а в эрлифтных биореакторах для аэрации и перемешивания используют газ (обычно воздух), который подается под давлением через разбрызгиватель в дне сосуда. При этом во всем объеме происходит непрерывная циркуляция жидкой среды. Барботажные колонны сходны с эрлифтными реакторами, но их недостатком является отсутствие циркуляции культуральной среды. Для обеспечения стерильности, постоянства pH, температуры и других параметров используют разные способы в зависимости от дизайна биореактора. Для синтеза рекомбинантных белков применяют двухступенчатые процессы ферментации, осуществляемые в тандемных эрлифтных биореакторах или в одном реакторе с механическим перемешиванием.
Если синтезированный продукт накапливается в клетках, то их осаждают из культуральной среды центрифугированием или фильтрацией, затем разрушают ферментативными, химическими или механическими методами и выделяют нужный продукт. Если синтезируемый продукт секретируется в культуральную среду, то процедура его выделения и очистки значительно упрощается.
ЛИТЕРАТУРА
Bailey J.E., D.F. Olis. 1977. Biochemical Engineering Fundamentais. McGraw-Hill, New York, N.Y.
Charles M. 1985. Fermentation scale-up: problems and possibilities. Trends BiolechnoL 3:134-139.
Datar R. 1986. Economics of primary separation steps in relation to fermentation and genetic engineering. Process Biochem, 21: 19—29.
Engler C.R. 1985. Disruption of microbial cells, p. 305-324. In C.L. Cooney. A.E. Humphrey, M. Moo-Young (ed.), Comprehensive Biotechnology, vol. 2. Pergamon Press, Oxford, United Kingdom.
Giorgio R.J-, J.J. Wu, 1986. Design of large scale containment facilities for recombinant DNA fermentations. Trends BiotechnoL 4: 60—65.
Gösset G., R. de Anda, N. Cruz, A. Marttinez, R. Quintero, F. Bolivar. 1993. Recombinant protein production in cultures of an Escherichia coli trp strain. Appl. Microbiol. Biotechnol. 39: 541-546.
Grund G., C.W. Robinson, B.R, Click. 1991. Croos-flow ultrafiltration of proteins, p. 69—83. In M.D. White, S. Reuveny, A. Shaffetman (ed.), Biologicalsfrom Recombinant Microorganisms and Animal Cells: Production and Recovery. Verlag Chemie, Weinhetm, Germany.
Hart R.A., P.M. Lester, D.H. Reifsnyder, J.R. Ogez, S.E. Builder. 1994. Large scale, in situ isolation of periplasmic IGF-f from E. coli. Bio/Technology 12: 113-117.
Kroner K.H. 1986. Cross-flow filtration in the downstream processing of enzymes: current status. Biotechnol. Forum 3:20—31.
Kroner, K.H., H. Nissinen, H. Zeigler. 1987. Improved dynamic filtration of microbial suspensions. Bio/Technology 5:921-926.
Lee S.V. 1996. High cell-density culture of Escherichia coli. Trends Biotechnol, 14:98—105.
McKillip E.R., A.S. Giles, M.H. Levner, P.P. Hung, R.N. Njorth. 1991. Bioreactors for large-scale t- PA production. Bio/Technology 9: 805-812.
Mendoza-Vega О., С. Hebert, S.W. Brown. 1994. Production of recombinant hirudin by high cell density fed-batch cultivations of a Saccharomyces
Промышленный синтез белков при участии рекомбинантных микроорганизмов 369
cerevisiae strain: physiological considerations during the bioprocess design. J. Biotechnol. 32:249-259.
Merchuk J.C. 1990. Why use airlift bioreactors? Trends Biotechnol. 8:66-71.
Park Т.Н., J.-H.Seo, H.C. Lim. 1991. Two-stage fermentation with bacteriophage λas an expression vector in Escherichia coli. Biotechnol. Bioeng. 37:297-302.
Paulson D.J., R.L. Wilson, D.D. Spatz. 1984. Cross-flow membrane technology and its applications. Food Technol. Dec. 1984: 77-87.
RamTrez D.M., W.F. Bentley. 1995. Fed-batch feeding and induction policies that improve foreign protein synthesis and stability by avoiding stress response. Bioiechnol. Bioeng. 471 596-608.
Reuss M. 1995. Stirred tank bioreactors, p. 207-255. In J.A. Asenjo, J. Merchuk (ed.), Bioreactor System Design. Marcel Dekker. Inc., New York, N.Y.
Robinson O.K., C.P. Chan, C.Vu Ip, P.K. Tsai, J. Tung, T.C. Seamans, A.B. Lenny, D.K. Lee, J. Irwin, M. Silberklang. 1994. Characterization of a recombinant antibody produced in the course of a high yield fed-batch process. Biotechnot. Bioeng. 44: 727-735.
Sauer T., C.W. Robinson, B.R. Click. 1989. Disruption of native and recombinant Escherichia coli in a high-pressure homogenizer, Biotechnot. Bioeng. 33: 1330-1342.
Sayadi S., M. Nasri, F. Berry, J.N. Barbotin, D. Thomas. 1987. Effect of temperature on the stability of plasmid pTG201 and productivity of xylE gene product in recombinant Escherichia coli: development of a two-stage chemostat with free and immobilized cells. J. Gen. Microbiol. 133: 1901-1908.
Schügerl K., A. Lübbert. 1995. Pneumatically agitated bioreactors, p. 257—303. In i.A. Asenjo, J. Merchuk (ed.), Bioreactor System Design. Marcel Dekker, Inc., New York, N.Y.
Schütte H., M.-R. Kula. 1990. Pilot- and process-scale techniques for cell disruption. Biotechnol. Appl. Biochem. 12: 599-620.
Seigel R., D.D.Y. Ryu. 1985. Kinetic study of instability of recombinant plasmid pPLc23f/pAl in E. coli using two-stage continuous culture system. Biotechnol. Bioeng. 27: 28-33.
Siegel M.H., H. Hallaille, J.C. Merchuk. 1988. Airlift reactors: design, operation, and applications. Adv. Biotechnol. Processes!: 79—124.
Strandberg L., K. Köhler, S.-O. Enfors. 1991. Large-scale fermentation and purification of a recombinant protein from Escherichia coli. Process Biochem. 26; 225-234.
Strandberg L., L. Andersson, S.-O. Enfors. 1994. The use of fed batch cultivation for achieving high cell densities in the production of a recombinant protein in Escherichia coli. F EM S Microbiol. Rev. 14: 53-56.
Strathman H. 1985. Membranes and membrane processes in biotechnology. Trends Biotechnol. 3: 112-118.
Tanny G.B., D. Mirelman, T. Pistole. 1980. Improved filtration techniques for concentrating and harvesting bacteria. Appl, Environ, Microbiol, 40: 269-273.
Tutunjian R.S. 1985. Scale-up considerations for membrane processes. Bio/Technology 3: 615-626.
Van Brunt J, 1985, Scale-up: the next hurdle. Bio/Technology 3: 419-424.
Van Brunt J. 1986. Fermentation economics. Bio/Technology 4: 395-401.
White M.D., B.R. Click, C.W. Robinson. 1995. Bacterial, yeast and fungal cultures: the effect of microorganism type and culture characteristics on bioreactor design and operation, p. 47—87. In J.A. Asenjo, J. Merchuk (ed.), Bioreactor System Design. Marcel Dekker, Inc., New York, N.Y.
Whitney, G.D., B.R. Click, C.W. Robinson. 1989. Induction of T4DNA ligase in a recombinant strain of Escherichia coli. Biotechnol. Bioeng, 33: 991-998.
Yamane T. 1995. Bioreactor operation modes, p. 479-509. In J.A. Asenjo and J. Merchuk (ed.), Bioreactor System Design. Marcel Dekker, Inc., New York, N.Y.
КОНТРОЛЬНЫЕ ВОПРОСЫ
1. В чем различия между периодической ферментацией, периодической ферментацией с добавлением субстрата и непрерывной ферментацией?
370 ГЛАВА 16
2. Какие параметры необходимо строго контролировать при оптимизации процесса ферментации?
3- Как влияет присутствие в клетке рекомби-нантной плазмиды на ее рост?
4. Как влияет перемешивание на доставку кислорода из культуральной среды к клеткам?
5. Для чего нужно стремиться максимально повысить плотность культуры при промышленной ферментации?
6. Каковы относительные преимущества и недостатки биореакторов с механическим перемешиванием и эрлифтных биореакторов?
7. Сравните процедуры выращивания и индукции культуры рекомбинантных микроорганизмов в двух тандемных биореакторах и в одном реакторе.
8. Какой обработке подвергают клеточную суспензию по завершении ферментации?
9. Какую стратегию вы бы выбрали для очистки рекомбинантного белка, секретируемого в культуральную среду?
10. Каковы преимущества и недостатки механического разрушения клеток по сравнению с химическим?
11. Как собирают клетки по завершении ферментации? Каковы преимущества и недостатки соответствующих методов?
ЧАСТЬ III.
Эукариотические системы
До недавнего времени высокопродуктивные сорта сельскохозяйственных растений и новые породы животных получали методом селекции. Однако этот подход, требующий для своей реализации много времени, уступил место методам, основанным на генной инженерии высших организмов. Теперь гены, обусловливающие специфические признаки, могут вводиться в клетки растений или животных и передаваться следующим поколениям (наследоваться). В ч. III мы рассмотрим, как получаются такие трансгенные растения и животные.
Уже создано несколько видов трансгенных растений, обладающих признаками, которые детерминируются генами, введенными в них генноинженерным и методами. К числу таких признаков относятся: способность синтезировать инсектициды; устойчивость к вирусным инфекциям и гербицидам; измененные сроки созревания плодов; измененная окраска цветков; повышенная пищевая ценность семян и самонесовместимость. Исследования трансгеноза у животных только начинаются, так что пока трудно предсказать, какие генетические признаки будут наследоваться видом-реципиентом. К настоящему времени выведены линии трансгенных мышей, которые используются как модельные системы для изучения механизма возникновения рака, муковисцидоза, болезни Альцгеймера и других заболеваний человека.
Технология рекомбинантных ДНК нашла широкое применение в изучении наследственных болезней человека и раз-
работке методов генной терапии. Так, используя специфический хромосомный сайт в качестве маркера, можно локализовать на хромосоме человека ген, ассоциированный с данным заболеванием, ничего не зная о механизме действия этого гена, а затем, используя клонированную последовательность, которая узнает этот маркерный сайт, попытаться идентифицировать дефектный ген. С помощью такого подхода уже были найдены и охарактеризованы гены некоторых болезней человека. Далее можно исследовать механизм действия нормального и дефектного генов и разработать эффективные методы лечения. В ч. III обсуждаются молекулярная генетика человека и генная терапия.