биология. EKZAMEN_BIO (Восстановлен). Краткий обзор 1) единство химического состава, 2) обмен веществ, 3) самовоспроизведение (репродукция), 4) наследственность
 Скачать 1.8 Mb. Скачать 1.8 Mb.
|

|
Химическая эволюция или пребиотическая эволюция — современная теория происхождения жизни — также опирается на идею самозарождения. Однако в основе ее лежит не внезапное (de novo) возникновение живых существ на Земле, а образование химических соединений и систем, которые составляют живую материю. Пребиотические вещества возникли из неорганических молекул под влиянием внешних энергетических и селекционных факторов и в силу развертывания процессов самоорганизации, свойственных всем относительно сложным системам, которыми, бесспорно, являются все углеродсодержащие молекулы. Она рассматривает химию древнейшей Земли, прежде всего химические реакции, протекавшие в примитивной атмосфере и в поверхностном слое воды, где, по всей вероятности, концентрировались легкие элементы, составляющие основу живой материи, и поглощалось огромное количество солнечной энергии. Гипотеза РНК-мира-гипотетический этап возникновения жизни на Земле С открытием генетических клеточных структур, с определением функции и строения ДНК, с развитием микробиологии появился новый кандидат на роль молекулы первожизни. Им стала молекула рибонуклеиновой кислоты – РНК. Эта макромолекула, входящая в состав всех живых клеток, представляет собой цепь нуклеотидов – простейших органических звеньев, состоящих из атомов азота, моносахарида – рибозы и фосфатной группы. Именно последовательность нуклеотидов является кодом наследственной информации. Уникальная способность некоторых молекул РНК вносить разрывы в другие цепочки или склеивать отдельные элементы РНК, а некоторые играют роль автокатализаторов – то есть способствуют быстрому самовоспроизведению. Некоторые из них имеют способность катализировать синтез белков, которые, в свою очередь, сами катализируют синтез РНК и свой собственный синтез (развитие трансляции). Некоторые молекулы РНК соединяются в двойную РНК-спираль, они развиваются в молекулы ДНК и носители наследственной информации (развитие транскрипции).Относительно небольшие размеры макромолекулы РНК и её упрощенное, по сравнению с ДНК, строение (в одну нить) сделали рибонуклеиновую кислоту главным кандидатом на роль основного элемента добиологических систем. Окончательно новую теорию возникновения живой материи на планете сформулировал в 1986 году Уолтер Гилберт, что молекулы РНК были предшественниками организмов. Важными каталитическими центрами рибосом являются РНК, а не, как раньше было принято, белки. Это показывает, что каталитическая функция РНК, как она предполагалась в гипотезе мира-РНК, используется сегодня живыми существами. Так как рибосомы считаются очень первоначальными клеточными органоидами, это открытие считается важным взносом в обоснование гипотезы мира-РНК. Уже можно уверенно сказать, что молекулы РНК могут из аминокислот синтезировать белки. Вопрос 141. «Горизонтальный перенос генов» Горизонтальный перенос генов — существующий в природе процесс, заключающийся в передаче генов между одновременно существующими взрослыми организмами (от одного генома к другому, в особенности между двумя видами), а не от родителей к потомству (вертикальный перенос генов). Примером может служить перенос генов устойчивости к ртути (mer-оперонов) между различными штаммами и видами природных бактерий. Горизонтальный перенос генов между различными видами организмов рассматривается в качестве одного из механизмов адаптивной эволюции. Искусственный горизонтальный перенос генов используется в генной инженерии. Горизонтальный перенос генов можно выявить по ряду показателей: 1. по нуклеотидному составу ДНК (ГЦ-содержание), который является видоспецифичным признаком. Отличие в нуклеотидном составе отдельного сегмента от остальной части генома является указанием на присутствие «чужих» генов; 2. по частоте встречаемости в гене определенных кодонов. В генах каждого вида преимущественно используется ограниченный набор кодонов. 3. на основании отличия в положении анализируемого гена на филогенетическом древе от большинства других генов. О «чужеродном» происхождении гена может говорить и высокая степень его сходства с гомологичным геном из отдаленного таксона при отсутствии подобного гена у филогенетически близких «родственников». 4. на основании сравнительного анализа белковых доменов. Большинство доменов характеризуются строго определенной функцией и представляют собой, таким образом, функциональные блоки белковых молекул. Возможны три варианта переносов: 1. Приобретение нового гена, для которого нет гомолога в собственном геноме и в геномах филогенетически родственных организмов. В этом случае возникает принципиально новое качество. 2. Приобретение паралогичного (структурно похожего) гена с генетически отдаленным родством. В результате такого переноса увеличивается функциональное разнообразие белков в клетке. 3. Приобретение нового гена ксенолога, функционально замещающего свой собственный ген, который при этом, как правило, элиминируется. Новый и старый гены структурно различаются между собой, но обеспечивают аналогичные физиологические функции. Горизонтально могут переноситься два типа последовательностей: 1) последовательности из транспозиционных элементов и 2) геномные последовательности Вопрос 142. Молекулярная филогенетика Молекулярная филогенетика — способ установления родственных связей между живыми организмами на основании изучения структуры полимерных макромолекул — ДНК, РНК и белков. Результатом молекулярно-филогенетического анализа является построение филогенетического дерева живых организмов. Близкое родство между живыми организмами обычно сопровождается большой степенью сходства в строении тех или иных макромолекул, а молекулы не родственных организмов сильно различаются между собой. Молекулярная филогения использует такие данные для построения филогенетического древа, которое отражает гипотетический ход эволюции исследуемых организмов. Вопрос 143. Генетический контроль метамерной организации организмов. Очевидно, что генетический контроль развития существует, ибо как тогда понять, почему из яйца крокодила развивается крокодил, а из яйца человека — человек. Каким образом гены определяют процесс развития? Это центральный и очень сложный вопрос, к которому ученые начинают подходить, но для всеобъемлющего и убедительного ответа на него данных явно недостаточно. Главным приемом ученых, изучающих генетику индивидуального развития, является использование мутаций. Выявив мутации, изменяющие онтогенез, исследователь проводит сравнение фенотипов мутантных особей с нормальными. Это помогает понять, как данный ген влияет на нормальное развитие. С помощью многочисленных сложных и остроумных методов стараются определить время и место действия гена. Анализ генетического контроля затрудняется несколькими моментами. Прежде всего тем, что роль генов неодинакова. Часть генома состоит из генов, определяющих так называемые жизненно важные функции и отвечающих, например, за синтез тРНК или ДНК-полимеразы, без которых невозможно функционирование ни одной клетки. Эти гены названы «house keeping» или генами «домашнего хозяйства». Другая часть генов непосредственно участвует в детерминации, дифференцировке и морфогенезе, т.е. функция их, по-видимому, более специфическая, ключевая. Для анализа генетического контроля необходимо, кроме того, знать место первичного действия данного гена, т.е. следует различать случаи относительной, или зависимой, плейотропии от прямой, или истинной, плейотропии. В случае относительной плейотропии, как, например, при серповидно-клеточной анемии, существует одно первичное место действия мутантного гена — гемоглобин в эритроцитах, а все остальные наблюдаемые при ней симптомы, такие, как нарушение умственной и физической деятельности, сердечная недостаточность, местные нарушения кровообращения, увеличение и фиброз селезенки и многие другие, возникают как следствие аномального гемоглобина. При прямой плейотропии все разнообразные дефекты, возникающие в различных тканях или органах, вызываются непосредственным действием одного и того же гена именно в этих разных местах. 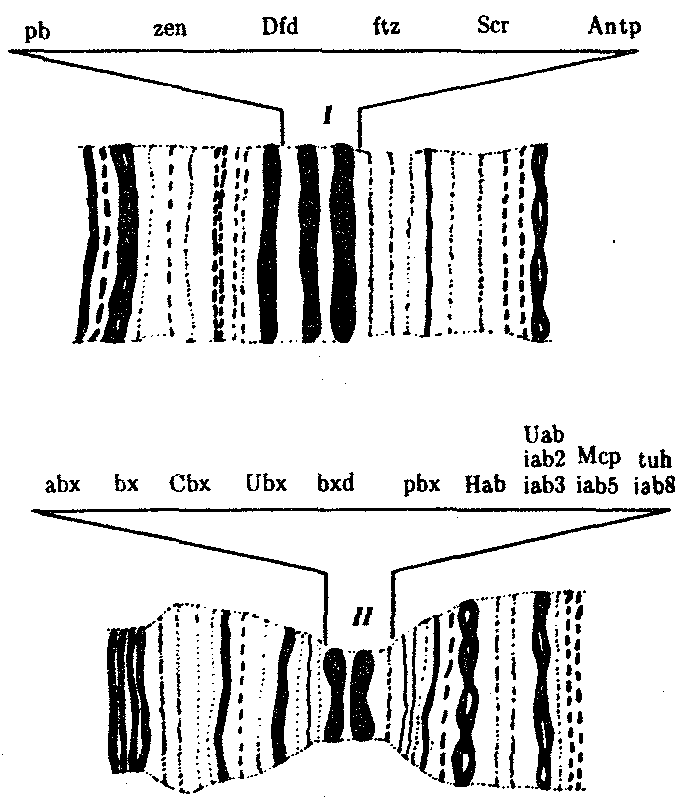 Рис. 8.14. Участки политенных хромосом дрозофилы с указанием расположения комплексов ANT-C(I) и ВХ-С(II) и генов, входящих в эти комплексы Наконец, следует различать еще два способа действия мутаций на фенотип, вызывающих дизруптивные либо гомеозисныеизменения. В первом случае, и это бывает чаще всего, мутации приводят к нарушению нормального развития, отсутствию или аномальному строению органов. В других случаях отклонение от нормы заключается в том, что под действием мутации типичный орган замещается гомологичным или совсем другим, но с нормальным строением. Это особый класс мутаций, описанный у насекомых и получивший название гомеозисных мутаций. Примером гомеозисных мутаций являются мутации в ВХ-С и ANT-C-комплексах генов у дрозофилы (рис. 8.14). ВХ-С (Bithorax Complex) и ANT-C (Antennapedia Complex) — это два набора генов, представляющих собой два кластера тесно сцепленных между собой генов, находящихся в одном плече 3-й хромосомы (рис. 8.15). В группу ВХ-С входит не менее 10 генов, которые отвечают за индивидуальность сегментов тела, расположенных сзади от среднегрудного сегмента. Чем больше дистальных локусов ВХ-С подвергается делении, тем больше брюшных сегментов превращается в грудные. При делении всего комплекса ВХ-С все сегменты от заднегрудного до 8-го брюшного превращаются в среднегрудь. В группу ANT-C входит не менее шести генов. Усиление в результате мутации функции этой группы вызывает превращение антенны в ногу, как должно быть в грудных сегментах. 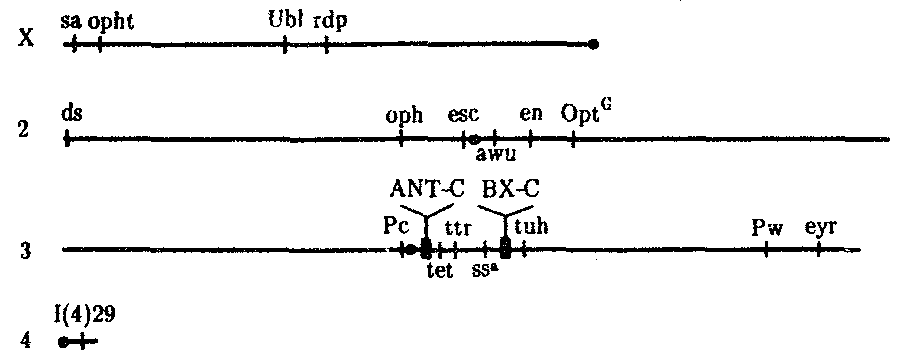 Рис. 8.15. Расположение гомеозисных комплексов мутантных генов ANT-C и ВХ-С в геноме дрозофилы: X, 2, 3, 4 —икс-, вторая, третья и четвертая хромосомы; черной точкой обозначены центромеры, латинскими буквами — некоторые гены Обширные генетические исследования гомеозисных мутаций у дрозофилы показали, что эти комплексы генов отвечают за направление развития сегментов тела, т.е. являются как бы морфогенетическими переключателями. Этот вывод имеет большое теоретическое значение, так как позволяет предположить наличие таких генов-переключателей направления развития и у других видов организмов. В предыдущих разделах были приведены примеры генетически запрограммированных процессов клеточного деления, клеточных миграций, клеточной гибели и клеточной дифференцировки, которые проявляются избирательно, т.е. в определенный период и в определенном месте. Можно привести еще множество примеров мутаций, которые показывают, как и когда действуют гены. Существуют мутации, которые указывают на существование у многих видов животных так называемых генов с материнским эффектом. Особенность этих генов состоит в том, что материнский геном во время овогенеза продуцирует ферменты, необходимые для метаболизма раннего зародыша, а также передает информацию, касающуюся расположения и организации структур зародыша, т.е. оказывает влияние на морфогенез. Поэтому самка, гомозиготная по рецессивному мутантному аллелю и продуцирующая аномальные яйца, даже при скрещивании с нормальным самцом дает нежизнеспособное потомство. Сама же она развивалась вполне нормально, поскольку ее мать в этом случае могла быть только гетерозиготной и в ее яйцах были все факторы, необходимые для раннего развития. Интересно, что если в дефектные яйца рецессивной самки ввести цитоплазму от нормальных яиц, то зародыши будут спасены. Факторы, детерминируемые генами с материнским эффектом, обычно оказывают свое влияние на зародыш до периода гаструляции. Начиная с гаструляции все большую роль играет информация самого зародыша. Известны и другие мутации, оказывающие влияние на раннее развитие, но не связанные с материнским эффектом. К ним относятся, например, мутации рибосомных генов. У шпорцевой лягушки (Xenopus laevis) мутанты были лишены части или всех рибосомных генов. Цитологически это проявлялось в полном или частичном отсутствии ядрышка. В гомозиготном состоянии мутантных аллелей зародыши становятся полностью нежизнеспособными на стадии выклева, так как у них не образуются новые рибосомы, а те, что были запасены в яйце, уже полностью использованы. У мышей также известен целый ряд рецессивных мутаций сложного локуса Т 17-й хромосомы, затрагивающих раннее развитие. Локус Т представлен множеством (117) аллелей, обозначаемых знаком t с дополнительными индексами: t1, t2, t3 и т.д. Около 30% t-генов в гомозиготном состоянии вызывает гибель зародышей, часть аллелей являются полулетальными. Весь этот ряд рецессивных аллелей t распадается на восемь групп, которые могут быть комплементарны друг другу и в гетерозиготном состоянии не приводить к гибели зародыша. Известны также и пять доминантных мутаций Т-локуса. Каждая из восьми групп обусловливает разного рода дефекты. Один из аллелей останавливает превращение морулы в бластоцисту, состоящую из трофобласта и эмбриобласта. Такие морулы гибнут. Другая мутация приводит к тому, что развившийся трофобласт не вступает в контакт со стенкой матки и зародыш тоже гибнет. Третьи мутантные зародыши не образуют внезародышевой эктодермы, у четвертых — гибнут клетки зародышевой эктодермы, у пятых — клетки зародышевой эктодермы не способны мигрировать в области первичной полоски и образовывать мезодерму, у шестых — уже образовавшиеся структуры нервной системы дегенерируют и т.д. Первичное нарушение, лежащее в основе всех этих эффектов, всего лишь одного локуса пока не выяснено. Однако очевидно, что локус Т играет первостепенную роль в морфогенезе эктодермы мышиного зародыша и организма в целом. Известны мутации с более специфическим действием. У аксолотля мутация с (cardial lethal), наследуемая как простой аутосомно-рецессивный признак, вызывает нарушение развития сердца и его неспособность к сокращениям. Мутантные особи раздуты переполняющей их жидкостью, а пищеварительная система и жабры у них недоразвиты. Установлено, что у позвоночных сердце развивается в результате индукционного воздействия со стороны головного участка энтодермы. Опыты по пересадке сердечной мезодермы показали, что мутантные зародыши не могут обеспечить развитие сердца из подсаженной нормальной сердечной мезодермы. Это указывает на отсутствие индукционной активности головного участка энтодермы у мутанта с генотипом cc. Неразрывную связь генов с морфогенезом отдельных органов и систем органов можно наблюдать также на примере мутации Tfm (Testicular feminization locus) у млекопитающих. У человека, мышей и крыс этот ген наследуется как сцепленный с полом. Самки, гетерозиготные по мутантному гену, т.е. ХTfmX, по существу, нормальны, но половина их генотипически мужских потомков ХTfmY имеют женский фенотип и стерильны. В основе этого дефекта лежит отсутствие специфического белка, служащего рецептором тестостерона. У особей ХTfmY рецептор не синтезируется. Самки ХTfmХTfm нормальны во всех отношениях и фертильны. Это свидетельствует о том, что нормальное половое развитие самки может проходить без продукта этого гена. Органогенез — период, когда действие мутаций проявляется в большой мере. Развитие каждого органа и тем более системы органов контролируется совокупным координированным действием сотен генов. У человека известны свыше 120 форм наследственной глухоты, которые возникают в результате экспрессии мутантных генов, отвечающих за формирование слухового анализатора. У человека описано также около 250 наследственных поражений глаз, около 150 наследственных аномалий развития скелета, не менее 18 генов, отвечающих за нормальную дифференциацию пола. О значении генетического контроля онтогенеза говорят многочисленные болезни, связанные с геномными и хромосомными мутациями. В целом генетический контроль онтогенеза очевиден, однако в процессе развития зародыш и его части обладают способностью к саморазвитию, регулируемому самой целостной развивающейся системой и не запрограммированному в генотипе зиготы. Вопрос 144. Характеристика доменов современного филогенетического древа. Филогенетическое дерево — дерево, отражающее эволюционные взаимосвязи между различными видами или другими сущностями, имеющими общего предка. Филогенетическое дерево показывает общее происхождение организмов из всех трёх доменов. Бактерии, эукариоты и археи. 1. Вершины филогенетического дерева делятся на три класса: листья, узлы и (максимум один) корень. Листья — это конечные вершины, то есть те, в которые входят ровно по одному ребру; каждый лист отображает некоторый вид живых организмов (или иной объект, подверженный эволюции, например, домен белка). Каждый узел представляет эволюционное событие: разделение предкового вида на два или более, которые в дальнейшем эволюционировали независимо. Корень представляет общего предка всех рассматриваемых объектов. Ребра филогенетического дерева принято называть «ветвями». 2. Идея «дерева» появилась в ранних взглядах на жизнь, как на процесс развития от простых форм к сложным. Современные эволюционные биологи продолжают использовать деревья для иллюстрации эволюции, так как они наглядно показывают взаимосвязи между живыми организмами. 3. Типы филогенетических деревьев. - Укоренённое дерево — дерево, содержащее выделенную вершину — корень. Укоренённое дерево можно считать ориентированным графом, поскольку на нём имеется естественная ориентация — от корня к листьям. Каждый узел укоренённого дерева отвечает последнему общему предку нижележащих листьев дерева. - Неукоренённое дерево не содержит корня и отражает связь листьев без предполагаемого положения общего предка. - Укоренённое и неукоренённое филогенетическое дерево может быть бифуркационным (эволюционные события состояли в происхождении от предкового объекта ровно двух потомков) или небифуркационным(к узлу могут подходить четыре и более ветви), а также маркированным(содержит названия листьев) или немаркированным(просто отражает топологию) 4. Филогения живых организмов(общее происхождение организмов всех трёх доменов):Бактерии; Артерии;Эукариот |