Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
 Скачать 7.39 Mb. Скачать 7.39 Mb.
|

|
Усиление секреции желудка и поджелудочной железы, типе;: трофия слизистой оболочки желудка, усиление моторики ж< лудка, тонкой кишки и желчного пузыря. Увеличение секреции бикарбонатов поджелудочной железо! торможение секреции соляной кислоты в желудке Усиление сокращения желчного пузыря и желчевыделени5 секреции ферментов поджелудочной железой, торможени секреции соляной кислоты в желудке, усиление в нем секр< ции пепсина, усиление моторики тонкой кишки Глюкозозависимое усиление высвобЪждения поджелудочно железой инсулина, торможение секреции и моторики ж< лудка, торможение'высвобождения гастрина Расслабление гладких мышц кровеносных сосудов (снижен артериального давления), желчного пузыря, сфинктеро торможение секреции желудка Усиление моторики желудка и Тонкой кишки, усиление секр< ции пепсина желудком, синтеза белка Антагонист холецистокинин-панкреозимина, усиливает прол! ферацию слизистой оболочки тонкой кишки, поджелудочнс железы и печени, участвует в регуляции обмана углеводе и липидов Торможение высвобождения желудочно-кишечных гормоне и секреции желез желудка , Стимуляция желудочной секреции через высвобожден! гастрина, усиление сокращений желчного пузыря и выдел ние ферментов поджелудочной железой через высвобо» дение ХЦК-ПЗ, усиление высвобождения энтероглюкаг.он нейротензина и ПП Торможение секреции ферментов поджелудочной железо усиление высвобождения гастрина. Торможение секреции соляной кислоты железами желудк усиление высвобождения глюкагона Усиление моторики кишечника, слюноотделения, торможен: высвобождения инсулина и всасывания натрия Стимуляция секреции поджелудочной железой фермен' химотрипсиногена Усиление моторики ворсинок тонкой кишки Большинство гормонов обладает широкими спектрами действия и высвобождаете из соответствующих клеток под влиянием нескольких факторов. ; Некоторые гастроинтестинальные пептид-гормоны (гастрин, холецистокинин-Панк- реозимйн, ВИН, вещество П, энкефалин, нейротензин, бомбезиноподобный пептид) обнаружены в различных структурах мозга. Полагают, что это результат эмбрионального происхождения эндокринных клеток из нейроэктодермы или прилегающей к ней ткани. Значение этих веществ в ЦНС точно не установлено. Возможно, что данные пептид-гормоны выполняют\в мозге медиаторную функцию! Доказанной она считается лишь для некоторых-пептидов (ВИП, соматостатин, энкефалин, вещество П, бомбезин) , высвобождающихся в окончаниях ряда вегетативных нервных волокон, иннервирующих желудочно-кишечный тракт. Такие волокна названы пептидергическими (по аналогии с адрен- и холинергическими). ■ ; Перечисленные пептиды могут продуцироваться в соме нейронов,. транспортироваться по аксону и высвобождаться при деполяризации его окончания. Здесь он оказывает-'(каК нейромедиатор) локальное действие и быстро разрушается: Пептид может синтезироваться и в эндокринных клетках, высвобождаться из них под влиянием ряда факторов и оказывать локальное влияние, диффундируя; через ;интерстйциальную жид- сость от эндокринной клетки к клетке-мишени. Такое явление называется паракри'нией. К шслу паракринных пептидов относят, например, ВИП, субстанцию II, соматостатин. И, гаконец, пептиды, высвобождаясь в кровоток, циркулируют вместе с кровью в организме л действуют как типичные гормоны. Время их действия ограничивается Несколькими минутами, после чего гормоны разрушаются в почках и печени и выводятся из организма. Для поддержания определенной концентрации пептид-гормонов в крови они должны щпрерывно выделяться в: кровоток клетками-продуцентами. ; Действие интестинальных гормонов (энтеринов) не ограничивается пищеваритель- юй системой. ■ ■ Так,, гормональные факторы двенадцатиперстной кишки "увеличивают общий энергетический обмен, снижают аппетит, изменяют активность желез внутренней секции и т. д. Таким образом, интестинальным гормонам принадлежит большая роль ie только в регуляции деятельности органов пищеварения, но и обмена веществ орга - [изма в целом. Секреторные и мышечные клетки желудочно-кишечного тракта изменяют уровень функциональной активности в;зависимости, от рефлекторных интеро- и экстерорецептор- !ых и гуморальных влияний, а также в зависимости от уровня кровоснабжения (рис. 169). Деханиз.мы регуляции взаимосвязаны и контролируют ход пищеварительного процесса ia протяжении всего желудочно-кишечного тракта. . Эфферентные нервные и гормональные воздействия на органы пищеварения вызы- ают, по И. II. Павлову, три типа эффектов: функциональные, сосудодвигательные и рофйческие. Первый состоит в изменении, той или иной функциональной активности леток, органа или систейы, Второй тип эффектов — изменение уровня их .кровоснаб- <ения, приведение его в соответствие с уровнем функциональной активности-органа, юльшинство нервных и гормональных стимуляторов органов пищеварения усиливает х кровоснабжение, что является важным условием поддержания, высокой функциональ- ой активности данных органов. Третий тип эффектов включает в себя разнообразные зменения трофики органов пищеварения: процессы синтеза в них пищеварительных сек- етов,. изменения числа секреторных клеток в пищеварительных железах и т. д. Например, астрин увеличивает.число париетальных клеток в железах желудка;: холецистокинин- анкреозймин — гландулоцитов в ацинусах поджелудочной железы, И. П. Павлов основал учение о фазах секреции пищеварительных желез. По меха- изму изменения секреции пищеварительных желез, выделяют две фазы: сдожнореф- екторную и нервно-химическую. Первая осуществляется с помощью условных и безуловных рефлексов, вторая — нейрогуморальными механизмами. Деление на эти . две 1азы в значительной мере условно, так как нервные и гуморальные механизмы взацмо- вязаны. Секреция начинается с «мозговой» фазы и осуществляется по типу условного вид, запах пищи и т.д.), безусловного (раздражение рецепторов.полости рта и пище- ода) рефлексов. О С П г ^ I о I — 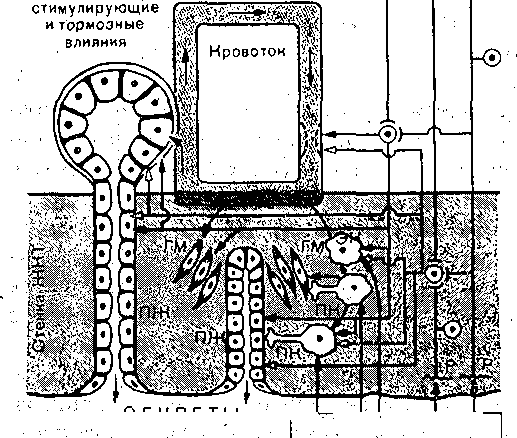 Энсторорецепторы ЦНС П ; Внешние По месту афферентных кейро-гуморальных воздействий:. 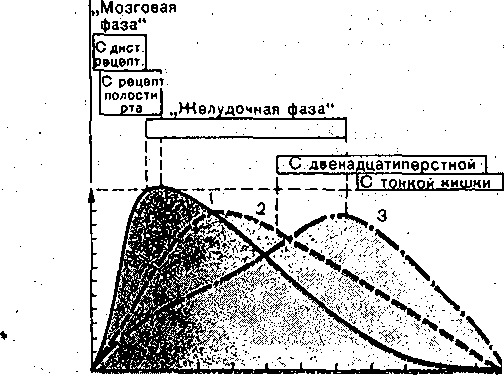 Корригирующие влияния Отимулирующие и " Полость желудочно иишечного тормозные влияния , тракта (ЖНТ) из полости ЩИТ По механизму воздействий: l^jffiljftj -слончкорефленторная фаза -ней рог у моральная фаза Рис. 169. Механизмы регуляции секреторной ■ - и моторной деятельности органов пищеваре- ,. Рис. 170, Фазы секреции г.кижых пище н,ия- . : .' . ■ • верительных желез. . Р — интерорецепторы, ЭК — эндокринные клетки, j __ желудочная секреция при выраженно продуцирующие гастроинтестинальные гормоны, «мозговой» фазе; 2 — при ее заторможее ПЖ — пищеварительные железы'; ГМ — гладкие ности; 3 — секреция' поджелудочной желе миоциты; эфферентные симпатические; (С) и парат ;ш симпатические (П) влияния ЦНС на желудочно- кишечный тракт (ЖКТ); ПК — паракринный путь . ; влияния. - ! ■■ ■;''■;'■ ■■.".-' - ■':''< Возбуждение-'секреции путем воздействий с желудка (раздражение его механ< йХеморецепторов; высвобождение гастрина) обозначается,желудочной фазой секреци; а посредством интестинальных: гормонов и раздражения рецепторов двенадцатиперстнс и тощей кишки - кишечной фазой секреции. По характеру влияний регуляторные механизмы можно разделить на пусковые и ко ригирующие. Роль последних особенно велика в обеспечении приспособления количесп и свойств пищеварительных Секретов к количеству и ряду свойств пищевого содержимо; желудка и кишечника. В месте действия раздражителя, и в каудальных отделах пищеварительного тракт всегда усиливается активность моторного и секреторного аппаратов, а в краниальнс расположенных отделах— торможение. Если пища в том или ином отделе желудочн» кишечного тракта недостаточно подверглась перевариванию, то в нем происходит з, держка пищевой массы и увеличивается секреция, что компенсирует начальное нед статочное переваривание пищи. Переход пищевых масс, недостаточно обработаннь секретами, в дистальнее расположенные отделы усиливает в них секреторные и гидрол. тические процессы. Так, при недостаточности желудочной секреции .в «мозговую» фа: удлиняется период секреции за счет ее желудочной и кишечной фаз (рис. 170). Так! образом, пищеварительный конвейер функционально составляет единое целое. Наход щаяся в пищеварительном тракте смесь пищевого содержимого с пищеваритеЛьныи соками является не только объектом пищеварительных воздействий. Параметры эт< смеси (консистенция, рН, осмотическое давление, концентрация ферментов и особен] промежуточных и конечных продуктов ферментного гидролиза питательных вещест есть средство регуляции самого пищеварительного процесса, они изменяют секреци моторику и всасывание в .желудочно-кишечном тракте. МЕТОДЫ ИЗУЧЕНИЯ ФУНКЦИЙ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА Основы современной физиологии пищеварения разработаны И. П. Павловым и его школой. До И. П. Павлова функции органов пищеварения изучали в основном в острых опытах, в которых нарушалось нормальное состояние организма вследствие наносимой >ис. 171. Эзофаготомиро ванна я собака с травмы. В 1842 г. московский хирург >истулой желудка (по , И. П. Павлову}. В. А. Басов предложил получать желудочное . содержимое посредством создания «искусственного хода в желудок». Эксперимен- ально-хирургическую методику исследования функции органов пищеварения в хронических ■ опытах И. П. Павлов довел до совершенства. Соблюдая все правила хирургии, производили операцию наложения фистулы того или иного отдела пищеварительного тракта. Фистулой называется искусственное сообщение полого органа или протока железы с нешней средой.). К опытам на таких животных приступали, когда они полностью оправятся от операми, рана заживет и органы пищеварения станут функционировать нормально. Как пра- ило, животные живут и участвуют в экспериментах длительное время. Чистый желудочный сок получают у животных с фистулой желудка и эзофаготомией i опыте т. н. «мнимого кормления» (операция предложена И. П. Павловым и Е. О. Щу- ювой-Симановской в 1899 г.). Животное с гастроэзофагостомой может, не насыщаясь, есть часами, так как пища/не поступает в желудок, а выпадает из отверстия ищевода наружу (рис. 171). При'таком мнимом кормлении из открытой желудочной жстулы изливается сок в большем количестве. Питание оперированных животных про- зводится' путем введения пищи и жидкостей в желудок через фистулу и отверстие в ищеводе. И. П. Павлов разработал операцию изолированного желудочка большой кривизны оставлением серозно-мышечного «мостика», со стороны кардии. В этом мостике прохо- ят сохраненные веточки блуждающего нерва, иннервирующего, изолированный желудо- ек, который адекватно отражает динамику секреторного процесса, в том числе иачаль- ые его рефлекторные фазы (рис. 172). И. П. Павловым предложена операция выведения в кожную рану общего желчного ротока, что дает возможность изучать механизмы выделения желчи. Исследование кишечной секреции производится на изолированных отрезках тонкой ишки (рис. 173). Один.(Тири) или оба (Тири — Велла) конца изолированного отрезка энкой кишки выводят в кожную рану. Ряд методов изоляции отрезка тонкой кишки редложен в павловской лаборатории. Для изучения моторной деятельности желудочно-кишечного тракта также широко ^пользуются животные с фистулами желудка и кишечника. Широко применяются рентгенологические методы, в том числе рентгенокинемато- зафия. , 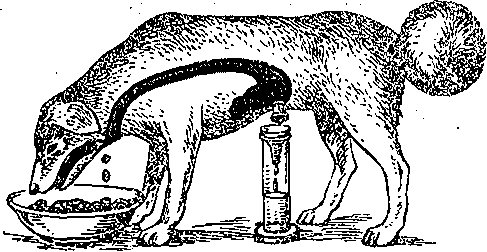 В экспериментах по изучению процесса всасывания используется метод забора крови, гтекающей от пищеварительных органов. Для этого к стенке сосудов пришивают трубки, входящие наружу. Через них, проколов сосуд иглой, можно получить кровь во время Фонического опыта (ангиостомическая методика Е. С- Лондона). Благодаря изобрете- ш полимерных катетеров и применению аитикоагулянтов в последнее время катетеры оживляют» в кровеносные сосуды и через них получают-кровь для анализа длительное )емя (вне опыта катетеры герметически закрываются) . Для исследования всасывания эжно собирать оттекающую от желудка и тонкой кишки лимфу, в которой определяются юсавшиеся вещества.
Рис! 172. Варианты операций изолированного желудочка (схема).
Рис. 173. Варианты изолированной петли тонкой кишки (схема). 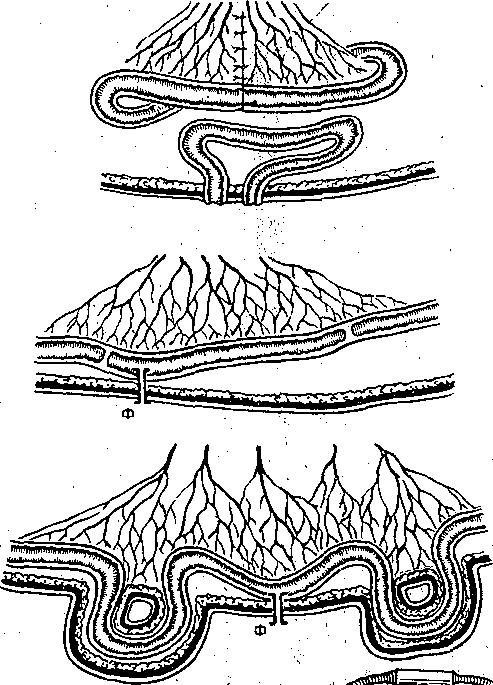 Энтероэнтероэнастомоз 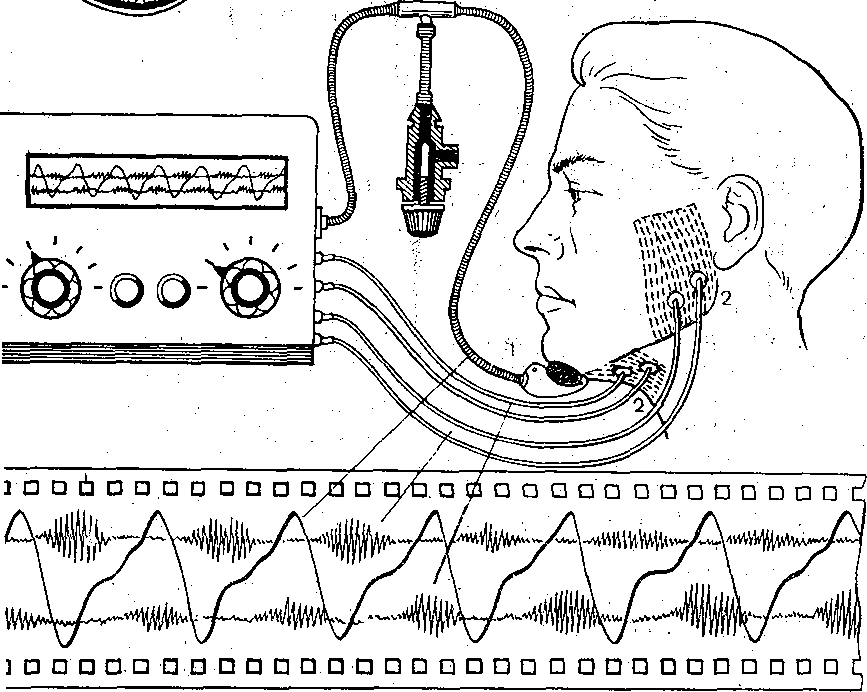 I — изолированная петля с выведенным в кожную рану дистальным ее концом; 2 — изолированная петля с выведенными наружу двумя "ее концами; 3 — петля с изолированной ее полостью; серозно — мышечный слой сохранен; 4 — в. кожные лоскуты выведены два участка тонкой кишки, пережатие их изолирует во время опыта полость кишечной петли с фистульной трубкой. (Ф). Рис. 174. Регистрация функции жевательной, мускулатуры методом мноэлектромастикациа- графии. (схема). 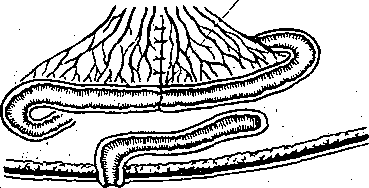 Эктероэнтероанастомоз 1 —датчик мастикациографа; 2 — электроды для отведения/биопотенциалов жевательных мышц. Рис. 175. Собирание слюны, у человека с помощью капсулы Лешли — Красногорского. Вид капсулы .изображен в нижней .Часта рисунка. 1 — Трубка для отсасывлния воздуха из внешней камеры капсулы; 2 — трубка для оттока слюны из внутренней камеры капсулы. . . Рис. 176. Зонд для определения кислотности разных зон желудка (по Линару). 1 — штепсель для соединения электродов, с ре- ги.стратором рН; 2 — резиновая трубка с отверстиями • (3), соединяющая резиновый баллон (6) с регистратором; 4 — резиновый зонд; 5 — провода; 7—8 — рН-оливы; 9 -• сурьмяные электроды; 10 - каломе.-евые электроды. Изучение деятельности пищеварительной системы ч е л о'в е к а требует специальных методических подходов. : Акт жевания исследуют путем регистрации движений нижней челюсти (мастикаци- ография) иногда с одновременной электромиографией жевательных мышц (рис. 174). При; помощи капсул Лешли — Красногорского (рис. 175) можно собрать раздельно слюну околоушной, подчелюстной и подъязычной желез. Изучение пищеварительных органов, расположённых в брюшной полости, требует иных методических приемов. Для учета с е к р е т о р н о й д е я т е л ь н о с т и Пищеварительных желез человека используются зондовые и беззондовые методы. В методах первой группы обследуемый проглатывает резиновую трубку (или вводят ее через нос), один конец которой достигает полости желудка или двенадцатиперстной Кишки (зонд может быть проведен й ниже), При помощи специального зонда определяют рН в желудке и верхних отделах кишечника (рис. 176). Зонд! может, иметь несколько датчиков рН, расположенных на различных уровнях.. Получаемые данные характеризуют кислото- выделительную деятельность, эвакуацию содержимого желудка в кишечник, транзит кислой жидкости Через двенадцатиперстную кишку и т.д. С развитием радиоэлектроники получила применение радиотелеметрическая методика. Сущность ее- заключается в том, что человеку дают проглотить миниатюрный радиопередатчик — радиопилюлю. Она! состоит из генератора электромагнитные колебаний, источника питания ..(сухой элемент, или аккумулятор) и датчика. Под влиянием воспринимаемых параметров датчик меняет частоту излучаемых радиопилюлей колеба- 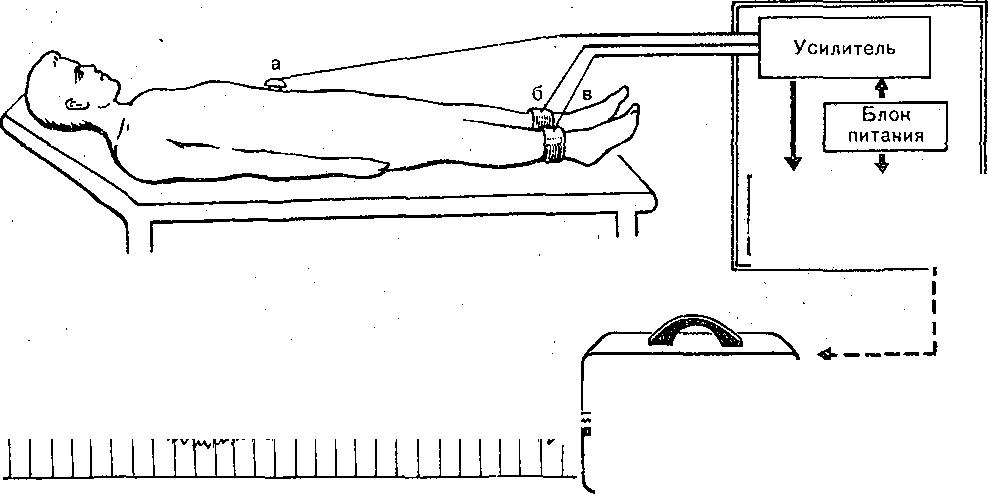 Блок—схема электрогастрографа ЭГС^З. Чернильно- пишущий I электромагнитный peJ гистратор и лентопрс* тяжный механизм | оооооооооо О'ОООйООО 00 00 ООО ООО о о Внешний вид электрогастрографа Элентрогастрограмма Рис. 177- Электрогастрография. а ,— активный электрод на коже в области желудка; б, в — индифферентные электроды на нижних конечностях (по М. В. Собакину). ний. Они воспринимаются антенной, надетой на обследуемого, и радиоприемником с записывающим устройством. Радиопилюля свободно проходит по желудочно-кишечному тракту. С ее помощью можно оценить секреторную деятельность желудка и моторную активность его и кишечника, а также гидролиз ряда питательных веществ. Моторную активность желудка можно изучать электрогастрографически, отводя с кожи живота и конечностей человека биопотенциалы (рис. 177), генерируемые гладкими мышцами желудка. Этот метод модернизируется для регистрации моторной активности •тонкой, толстой кишки и желчного пузыря. Для изучения моторной активности органов пищеварения человека широкое применение нашли рентгенологические методы (рентгеноскопия, рентгенография, рентгено- кинематография). Следует отметить также методы эндоскопии, которые позволяют осмотреть, сфотографировать слизистую оболочку желудка и начального отдела кишечника, устья выводных протоков, взять для исследования маленький кусочек слизистой оболочки (биопсия). Взятые кусочки подвергают гистологическому и биохимическому исследованию. . Для характеристики гидролиза белков и всасывания аминокислот широкое распространение получили пробы с мечеными белками (казеином или альбумином). Гидролиз и всасывание жиров исследуют также методами с дачей обследуемым меченых и немеченых жиров с последующим динамическим учетом содержания хило- микр,онов (микрокапельки жира, окруженные липопротеидной мембраной) или общих липидов в сыворотке крови. Широко применяется учет жира в кале. Существует и ряд других методов, подробно описанных в специальных курсах. ПИЩЕВАРЕНИЕ В ПОЛОСТИ РТА Переработка принятой пи-щи начинается в полости рта. Здесь происходят ее измельчение tсмачивание слюной, анализ вкусовых свойств пищи, начальный гидролиз некоторых пищевых веществ и формирование пищевого комка. Средняя длительность пребывания пищи в полости рта 15—18 с. II III V iV Рис. 178. Кимограмма жевательного периода (по Рубинову). Фазы: I — покоя; II — введения пиши в рот; III — ориентировочная; {V — основная; V — формирования пищевого комка и. проглатывания пиши,, внизу — отметка времени Г с, Поступившая в рот пища раздражает вкусовые, тактильные и температурные рецепторы. Вкусовые рецепторы расположены преимущественно в сосочках языка и рассеяны в слизистой оболочке.полости рта, тактильк-ые, температурные и болевые — по всей слизистой оболочке. Сигналы от этих рецепторов п-о центростремительным нервным волокнам тройничного, лицевого и языкоглоточного нервов доходят до нервных центров ряда рефлексов. Центробежные импульсы от этих центров рефлекторно возбуждают секрецию слюнных, желудочных и поджелудочной желез, выход желчи в двенадцатиперстную кишку, изменяют моторную активность желудка. Раздражение рецепторов полости рта имеет; важное значение в осуществлении актов жевания и глотания. Таким образом, несмотря на то что пребывание пищи во рту кратковременно, этот отдел пищеварительного тракта оказывает влияние на все этапы переработки пиши.- Жевание. В полость рта пища поступает в виде кусков, смесей разного состава и консистенции или жидкостей. В зависимости от этого пища или сразу проглатывается, или подвергается предварительной механической и химической обработке. Акт жевания совершается рефлекторно. Находящаяся во рту пища раздражает рецепторы, - от них сигналы по афферентным волокнам тройничного нерва передаются в центр жевания, а от него по эфферентным волокнам тройничного нерва — к жевательным мышцам. В координации акта жевания имеют также важное значение сигналы от проприорецепторов жевательных мышц, Жевательный период имеет фазы различной длительности в зависимости от свойств пережевываемой пищи (рис. 178) (покой, введение пищи в рот, ориентировочная, основная; формирование пищевого комка и глотание). Слюноотделение. На начальном этапе пищеварения, велика, роль слюны. Она продуцируется тремя парами крупных слюнных желез: околоушными, подчелюстными и подъязычными — и множеством мелких железок, находящихся на поверхности' языка, в слизистой оболочке неба и щек. Из желез по выводным протокам слюна поступает в полость рта. В зависимости от вырабатываемого секрета слюнные -железы бывают трех типов: серозные (вырабатывают жидкий секрет, не содержащий слизи — муцина); смешанные (вырабатывают серозно-слизистый секрет) и слизистые (вырабатывают слюну, богатую муцином) . Околоушная железа и малые железы боковых поверхностей языка имеют серозные клетки и продуцируют жидкую слюну. Слизистые железы расположены на корне языка и неба. В подчелюстной и подъязычной железах имеются серозные и слизистые клетки,-поэтому их называют смешанными. Смешанные железы находятся также в слизистой оболочке кончика языка, щек, губ. Из ацинусов слюнных желез секрет поступает в систему укрупняющихся протоков, собирающихся в общий выводной проток, выносящий слюну в полость рта. Вне приема пищи .у человека слюна выделяется в среднем 0,24 мл/мин для увлажнения полости рта, при жевании —. 3—3,5 мл/мин (около 200 мл/ч) в зависимости от вида.принимаемой пищи. В ответ на вводимый раствор лимонной кислоты слюноотделение может достигать 7,4 мл/мин. За сутки продуцируется 0,5—2,0 л слюны, около трети ее образуется околоушными железами. Состав и свойства слюны. Слюна представляет собой вязкую слегка опалесцирую- щую и мутноватую жидкость с плотностью 1,001 — 1,017. Состав слюны в большей мере зависит От скорости ее секрециц; рН смешанной слюны 5,8—7,4; рН слюны околоушных желез ниже (5,81), чем подчелюстных (6,39). С увеличением скорости секреции рН повышается до 7,8. ' -• Смешанная слюна содержит 99,4—99,5% воды, остальное — сухой остаток. Неорганические компоненты слюны: хлориды и карбонаты, фосфаты и другие соли натрия, калия, кальция, магния и др. Концентрация электролитов неодинакова в слюне разных слюнных желез и увеличивается с повышением скорости ее секреции, но слюна и в этих условиях имеет более низкое осмотическое Давление, чем плазма Крови. Слюна содержит органические вещества, которых в 2—3 раза больше, чем минеральных солей. Органические вещества являются продуктом секреторной деятельности слюнных желез, обмена веществ в них и частично транспортируются из крови. В составе слюны выделяются различные белки, свободные аминокислоты, некоторые углеводы, мочевина, аммиак, креатинин и другие вещества. Слюна содержит муцин, котбрый- придает ей вязкость, благодаря наличию муцина пропитанный слюной пищевой комок легко проглатывается. , Слюна достаточно богата ферментами, хотя содержание некоторых из них невелико. Слюна Человека Обладает Способностью активно гидролизовать углеводы. Это осуществляется а-амилазой, расщепляющей полисахариды (крахмал, гликоген) с образованием декстринов, а затем дисахаров (мальтозы) и частично глюкозы. Дисахаридазная активность слюны низкая. Амилаза слюны, начинает свое действие в полости'рта, но оно незначительно вследствие кратковременного пребывания здесь пищи. Гидролиз углеводов ферментами слюны продолжается в желудке, пока в глубокие слои его пищевого содержимого не проникает кислый желудочный сок, прекращающий действие карбогидраз и инактивирующий их. , В слюне содержится ряд других ферментов: протеиназы (катепсины, саливаин, глан- дулаин), липазы, щелочная и кислая фосфатазы, РНК-азы. Не исключено, что и они принимают участие в процессе пищеварения, но активность их невелика. Слюна обладает бактерицидным свойством за счет содержащегося в ней фермента лизоцима (мурамиДаза) . В слюне содержится калликреин, который принимает участие в образовании кининов, расширяющих кровеносные"сосуды, что может иметь значение в увеличении кровоснабжения слюнных (и других) желез при приеме ниши. Ферментный состав и свойства слюны изменяются с возрастом человека, зависят от режима питания и вида пищи. На пищевые вещества выделяется более вязкая слюна и тем больше, чем суше принимаемая пища. На отвергаемые вещества, кислоты, Горечи выделяется значительное количество более жидкой слюны. Адаптация слюноотделения к виду принимаемой пищи выражается не только В изменении объёма и вязкости слюны, но и ее каталитической активности. Количество й состав слюны в связи с приемом пиЩи определяются регуляторными воздействиями на слюнные железы. ' Регуляция слюноотделения Прием пищи возбуждает слюноотделение рефлекторно. При достаточно сильном раздражении и высокой возбудимости пищевого центра слюноотделение начинается через 1—3 с, а при слабой силе раздражения латентный период Достигает 20—30 с. Слюноотделение продолжается бесь период еды и прекращается вскоре после нее. : •'■■" , От рецепторов полости рта сигналы передаются в ЦНС но афферентным волокнам тройничного, лицевого, языкоглоточного и блуждающего нервов. Основной слюноотделительный центр расположен в продолговатом мозге: Именно сюда, а также в боковые рога верхних грудных сегментов спинного мозга поступают сигналы из полости рта' и расположенных выше отделов мозга. Отсюда влияния но эфферентным парасимпатическим и; симпатическим нервным волокнам направляются к слюнным железам. Парасимпатическая иннервация слюнных желез начинается из ядер продолговатого мозга (рис. 179). Симпатическая иннервация слюнных желез осуществляется от боковых рогов II—IV. грудных сегментов спинного' мозга. ' ' а - ' б' ^" Рис. 179. Схема путей рефлекторного возбуждения подчелюстцой (а) и околоушной (б) слюнных желез. .:■ ■;.'.... ...-. .; ... .. ■ а: I — тройничный иёра;, 2 узел тройничного нерва -(Гассеров); 3 — ядро лицевого нерва; 4 — лицевой нерв; 5 — коленчатый узел; 6- барабанная струна; 7 - язычный керв;. 8 — подчелюстная железа и подчелюстной ганглий; 9 крылонебныЙ ганглий; 10 - центр слюноотделения, б: I — тройничный узел; 2 - тройничный нерв; 3 — ядро язЫ ко глоточного нерва; 4 - лицевой нерв; 5 •• коленчатый узел; 6 — верхнечелюстной нерв;.7 — ушной узел; 8 - ушно-височный нерв; 9 — б&р.абаннаяструна; 10 — язычный нерв; 11 —• пещеристый узел; 12 — язы ко глоточный' нерв; 13 — околоушная слюнная железа. Выделены чувствительные нервные волокна, идущие о г ре цеп тор ных аппаратов языка, и преганглионарные парасимпатические волокна; пунктирная линия—.постганглионарные парасимпатические волокна. В окончаниях постганглионарных парасимпатических волокон высвобождается медиатор — ацетилхолин, возбуждающий секреторные клетки слюнных желез.. При этом выделяете я бол ьшое количе ство жидкой слюны. Дл ительное раздражение парасимпат и - ческих'нервов вызывает истощение слюнных желез: снижает с я соде р ж а н и е в слюне органических компонентов, п гландулоцитах резко уменьшается количество гранул. В окончаниях симпатических нервных волокон высвобождается норадреналин. Под его влиянием выделяется небольшое количество густой слюны. Раздражение симпатических нервов усиливает образование гранул секрета в гландулоцитах. Одновременное или предшествующее раздражение парасимпатических нервов усиливает симпатические секреторные эффекты. Ответная реакция при раздражении симпатических нервов возникает Спустя более длительный латентный период, чем После раздражения парасимпатических. ■.-,'■■■"■: : ; .Различия в секреторной деятельности слюнных желез в ответ на прием различных видов пищи можно объяснить модуляцией частоты импульсов по парасимпатическим и симпатическим нервным волокнам и изменением соотношений между их влияниями. Усиление слюноотделения сопровождается расширением кровеносных сосудов железы, что: повышает уровень ее кровоснабжения. Сосудорасширяющие эффекты в железе вызываются парасимпатическими нервными волокнами, образующимися в ней продуктами обмена веществ, а также кининами. Полагают также, что определенную роль, играет уменьшение сопротивления сосудов в результате освобождения железы от некоторого количества секрета. •л . Слюноотделениеначинается по типу условных рефлексов — в ответ на вид и запах пищи. Рефлекторные влияния могут и тормозить слюноотделение, вплоть до его прекращения. Такое торможение может быть вызвано болевым раздражением, отрицательными эмоциями, умственным напряжением, дегидратацией организма. Все эти воздействия снижают активность пищевого центра и его части — центра слюноотделения. Возбудителями последнего могут быть некоторые гуморальные вещества. Так, обильное отделение слюны наблюдаетхя при асфиксии вследствие раздражения центра слюноотделения угольной кислотой. Раздражают его некоторые токсины и при этом наблюдается обильное слюноотделение. , . Перерезка парасимпатических нервов, иннервирующих слюнные железы, временно резко усиливает выделение ими слюны — паралитическая секреция. В первые три дня непрерывное слюноотделение обусловлено повышенным выделением ацетилхолина вследствие дегенерации нейронов, поэтому эта секреция называется дегенеративной. В ^последующие дни паралитическая секреция связана с повышенной чувствительностью денервированной железы к ряду веществ, циркулирующих в крови, к которым железа с нормальной.иннервацией малочувствительна. Холиномиметические фармакологические вещества вызывают усиление слюноотделения, холинолитические — тормозят его. Глотание Глотание является рефлекторным актом, центр его находится в продолговатом мозге, на дне IV желудочка. Доказательством рефлекторной природы глотания служит следующее: если смазать корень языка и глотки раствором кокаина и выключить таким образом рецепторы их слизистой оболочки, то глотание осуществляться не будет. Глотание невозможно и после перерезки эфферентных нервов глотки. ' Рефлекс глотания состоит из ряда последовательных звеньев. Строгая координация звеньев глотательного рефлекса обеспечивается сложными взаимосвязями различных отделов ЦНС — от продолговатого мозга до коры большого мозга. Центр глотания связан с центром дыхания, что обеспечивает задержку дыхания во время глотания и имеет значение в предотвращении попадания пищи в воздухоносные пути. Связь центра глотания с центром регуляции сердечной деятельности подтверждается учащением сердечных сокращений во время глотания. Акт глотания делится на три фазы: I) ротовую (произвольную), 2) глоточную (быструю непроизвольную),^) пищеводную (медленную непроизвольную). Акт глотания изучен рентгеновским методом (рис. 180). : Из пищевой пережёванной массы во рту отделяется пищевой комок объемом 5—15 см3, который движениями языка и щек перемещается на спинку языка. Сокращениями передней части языка пшцевой комок прижимается к твердому небу, затем последовательными сокращениями средней части отжимается кзади'и переводится на корень языка за передние дужки (1-я фаза). Раздражение рецепторов слизистой оболочки корня языка рефлекторно вызывает сокращение мышд, приподнимающих мягкое небо, и' мышц языка, что препятствует попаданию пищи в полость носа. Движения языка способствуют проталкиванию пищи в полость глотки. Одновременно с этим происходит сокращение мышц, смещающих подъязычную кость и вызывающих поднятие гортани, вследствие чего закрывается вход в дыхательные пути, что препятствует поступлению в них пищи. Переводу пищи в глотку способствует повышение давления в полости рта. Возвращению пищи из глотки в полость рта препятствуют поднявшийся вверх корень языка и плотно прилегающие к нему дужки. Как только пища поступила в полость глотки, начинают сокращаться мышцы, суживающие просвет глотки выше пищевого комка, вследствие чего последний передвигается в пищевод. Существенное значение при этом имеет градиент давления между полостью глотки и началом пищевода. Перёд глотанием глоточно-пищеводный сфинктер закрыт, во время глотания давление в глотке повышается до 6 Па (45 мм рт. ст.) и через открывшийся сфинктер пищевой комок поступает в цищевод (2-я фаза), где давление в это время не более 4 Па (30 мм рт. ст.). Давление в глотке значительно выше, сфинктер закрыт и невозможно обратное забрасывание пищевого комка в глотку. Две фазы глотательного цикла длятся около 1с.  Рис. 180. Последовательность движении в полости рта и глотке при глотании. Вторую фазу глотания нельзя выполнить произвольно, если в полости рта нет пиши или слюны. В этом легко убедиться, делая подряд несколько глотательных движений: первое движение происходит легко, так как в полости рта всегда имеется небольшое количество слюны; при отсутствии слюны, которая проглочена, дальнейшие глотательные движения осуществить не удается. Третью фазу глотания составляют прохождение пищи по пищеводу и перевод ее в желудок. Движения пищевода взаимосвязаны с движениями глотательного аппарата и вызываются рефлекторно при каждом глотательном акте. Если в эксперименте на собаках перерезать пищевбд и вкладывать пищу непосредственно в него, то она передвигается в сторону желудка только после совершения глотательных движений. Продолжительность продвижения по пищеводу твердой пйщи в среднем 8—9 с, жидкой 1.-2 с. В момент акта глотания пищевод подтягивается к зеву и начальная часть его. расширяется, принимая пищевой комок. . Сокращения пищевода имеют характер волны, возникающей в верхней его части и распространяющейся в сторону желудка. При этом последовательно сокращаются кольцеобразно расположенные мышцы пищевода (в верхней трети поперечнополосатые, в нижних двух третях — гладкие), передвигая перед волной сокращения находящийся в несколько расширенной части пищевода пищевой комок в сторону желудка. Такой тип сокращений называется перистальтическим. Первичная перистальтическая волна, вызываемая актом глотания, Доходит до уровня пересечения пищевода с дугой аорты. Далее возникает вторичная перистальтическая волна, вызываемая не актом глотания, а первичной перистальтической волной. Вторичная волна продвигает пищевой комок до кардиальной части желудка. Средняя скорость распространения перистальтической волны по пищеводу человека 2—5 см/с, она в большой мере зависит от свойств пищи. Вторичная перистальтическая волна может быть вызвана остатком пищевого комка в нижней трети пищевода, благодаря чему этот остаток Переводится в желудок. Регуляция моторной деятельности пищевода осуществляется в основном эфферентными волокнами блуждающего и симпатического нервои; определенная роль'принадлежит интрамуральиым нервным образованиям пищевода. Вне глотательных движений вход из пищевода в желудок закрыт. КогДа пища й перистальтическая волна достигают конечной части пищевода, происходит рефлекторное понижение тонуса мышц кардии и пищевой комок переходит в желудок. При наполнении желудка тонус кардии повышается, что предупреждает забрасывание содержимого желудка в пищевод. Парасимпатические волокна блуждающего нерва стимулируют перистальтику пищевода и расслабляют кардию, симпатические волокна тормозят моторику пищевода, но повышают тонус кардиальной части: ПИЩЕВАРЕНИЕ В ЖЕЛУДКЕ Желудок принимает участие в пищеварительном конвейере, а также в межуточном обмене веществ, поддержании постоянства рН крови и-кроветворении (вырабатывает желудочный мукопротеид, или внутренний фактор Каела). 11ищеварительными функциями желудка являются: депонирование пищи, ее механическая и химическая обработка, постепенная порционная эвакуация пищевого содержимого в кишечник. Находясь в желудке в течение нескольких часов, пища набухает, разжижается, многие ее компоненты растворяются и подвергаются гидролизу ферментами слюны и желудочного сока. Желудочный сок обладает также антибактериальным действием. Карбогидразы, слюны продолжают действовать на углеводы пищи, находящиеся в центральной части пйщевого содержимого желудка (куда еще не диффундировал кислый желудочный сок, прекращающий действие карбогидраз слюны). Ферменты кислого желудочного сока воздействуют на пищевые белКи в относительно узкой зоне пищевого содержимого, находящегося в непосредственном контакте со слизистой оболочкой желудка и в небольшом удалении от нее, куда диффундировал желудочный сок и не был нейтрализован за счет буферных свойств пищи. Ширина этой зоны зависит от количества и свойств желудочного сока и принятой пищи. Вся масса пищи в желудке не смешивается с соком. По мере разжижения и химической обработки пищи ее слой, прилегающий к слизистой оболочке, движениями-желудка перемещается в его антральную часть, откуда эвакуируется в кишечник. Таким образом, пищеварение в желудке (по типу полостного) осуществляется некоторое время за счет слюны, но ведущее значение имеют секреторная и моторная деятельность самого желудка. ■ Секреторная деятельность желудка. |