Метод Физиология и анатомия как науки определение, основные понятия, значение в системе медицинских знаний. Значение знаний по физиологии и анатомии человека для врачафармацевта
 Скачать 0.97 Mb. Скачать 0.97 Mb.
|

|
Тазобедренный сустав (articulatio coxae, лат.) чашеобразный по форме. Образован головкой бедра и вертлужной впадиной тазовой кости, глубина которой увеличена за счет расположенной по ее краю хрящевой губы. Внутри сустава имеется круглая связка головки бедра, в которой проходят кровеносные сосуды и нервы, питающие головку. Суставная капсула прочная, укреплена связками. Наиболее крепкой из связок является расположенная спереди подвздошно-бедренная связка, выдерживающая нагрузку до 300 кг. Своим натяжением она препятствует разгибанию бедра. Коленный сустав (articulatio genus, лат.) — самый большой сложный сустав, по форме относится к мыщелковым суставам (рис. 2.25). В нем сочленяются суставные поверхности трех костей: мыщелков бедренной и большеберцовой кости и надколенника. Основное движение — сгибание и разгибание — совершается вокруг фронтальной оси. Суставная сумка укреплена прочными боковыми связками. Внутри сустава имеются хрящевые прокладки — латеральный и медиальный мениски, а также передняя и задняя крестообразные связки. Синовиальная оболочка суставной сумки образует завороты и складки. При согнутом колене боковые связки расслабляются, а на суставной поверхности большеберцовой кости оказываются задние отделы мы-щелков бедренной кости, имеющие шаровидную форму. В этом положении возможно незначительное вращение голени наружу и внутрь вокруг вертикальной оси. Вокруг сустава залегают синовиальные сумки; некоторые из них сообщаются с полостью коленного сустава. Соединение костей голени, между собой: верхние концы большеберцовой и малоберцовой костей образуют плоский малоподвижный сустав; тела этих костей соединены межкостной перепонкой, а нижние концы — связками. Голеностопный сустав (articulatio talocruralis, лат.) обеспечивает соединение стопы с голенью. По форме он блоковидный, образован нижними концами большеберцовой и малоберцовой костей, лодыжки которых в виде вилки охватывают таранную кость. Суставная сумка укреплена боковыми связками. В суставе возможно подошвенное сгибание и разгибание (или тыльное сгибание) вокруг фронтальной оси, проходящей через блок таранной кости. Кроме того, в состоянии подошвенного сгибания возможны небольшие боковые движения. Кости предплюсны соединяются суставами, сумки которых укреплены короткими связками как с подошвенной, так и с тыльной стороны стопы. Движения в этих суставах дополняют движения в вышележащем голеностопном суставе, чем достигается большая свобода движений всей стопы. Из суставов предплюсны имеют практическое значение таранно-ладьевидный и пяточно-кубовидный, которые соединяются в хирургии в один поперечный сустав предплюсны (сустав Шопара). Кости предплюсны соединяются с основаниями плюсневых костей плоскими малоподвижными предплюсна-плюсневыми суставами. Суставные капсулы их укреплены тыльными, подошвенными и межкостными связками. Плюснефаланговые и межфаланговые суставы по своему строению и движениям соответствуют аналогичным суставам кисти. 12. Типы соединения костей. Строение сустава. Классификация суставов. Соединения костей Каждая кость занимает в теле человека определенное место и всегда находится в непосредственной связи с другими костями, тесно прилегая к одной или нескольким костям. Различают два основных вида соединений костей: 1) непрерывные соединения — синартрозы, когда кости связаны одна с другой с помощью прокладки между ними из соединительной, хрящевой или костной ткани; 2) прерывные соединения — диартрозы (суставы), когда между сочленяющимися костями находится суставная полость и они удерживаются одна около другой с помощью замкнутой суставной капсулы и подкрепляющих ее связок и мышц. Переходной формой от непрерывных соединений к прерывным является полусустав — гемиартроз. Например, лобковый симфиз — это хрящевое соединение лобковых костей, внутри которого имеется небольшая щелевидная полость. К непрерывным соединениям относятся синдесмозы, синхондрозы и синостозы. По функции это или малоподвижные, или неподвижные соединения. Синдесмоз — соединительнотканное соединение костей. Самым распространенным видом синдесмоза являются связки (например, межостистые и межпоперечные связки позвоночника и т. д.). Некоторые связки имеют вид перепонок или мембран (мембраны между костями предплечья, голени). Разновидностью синдесмоза являются швы черепа. Сюда же относится форма укрепления зубов в луночках челюсти — вколачивание (гомфозис). Синхондроз — непрерывное соединение костей с помощью хряща, например соединение тел позвонков с помощью межпозвоночных хрящевых дисков. Синостоз — соединение костей с помощью костной ткани, костное сращение. Как правило, оно возникает на почве синхондроза (например, синостоз между телами затылочной и основной костей. К прерывным соединениям относятся суставы или сочленения (articulations, лат. - сустав). По функции это подвижные соединения. В суставе различают суставные поверхности сочленяющихся костей, окружающую их суставную капсулу и суставную полость. Суставные поверхности костей покрыты гладким и блестящим гиалиновым хрящом. Толщина суставного хряща колеблется от 0,5 до 4 мм. Суставная капсула имеет два слоя: наружный — фиброзный и сращенный с ним внутренний — синовиальный. Синовиальный слой образует складки, а в некоторых суставах — выпячивания, сумки (бурсы). Синовиальные сумки могут сообщаться с полостью сустава или быть изолированными. Располагаясь снаружи вокруг сустава в виде мягких прокладок между костью и сухожилиями мышц, они уменьшают трение. Внутренняя поверхность капсулы и суставные хрящи покрыты тонким слоем прозрачной тягучей жидкости — синовии, выделяемой клетками ворсинок синовиального слоя. Она выполняет роль смазки — уменьшает трение и способствует скольжению. Полость сустава представляет собой щелевидное пространство, ограниченное сочленяющимися поверхностями костей и суставной капсулой. Благодаря полному соответствию рельефа суставных хрящей и отрицательному давлению внутри сустава суставные поверхности костей всегда плотно прилежат друг к другу. Этому способствуют также связочный аппарат, укрепляющий суставную капсулу снаружи, и тяга мышц. Связки и сухожилия мышц составляют вспомогательный аппарат сустава. Одни связки укрепляют капсулу в местах наибольшего ее натяжения и ограничивают движение. Это тормозящие связки. Другие способствуют движению сустава в определенном направлении. Это направляющие связки. В результате неудачного движения или травмы может произойти растяжение и даже разрыв связок, следствием чего бывает смещение костей в суставе — вывих. К добавочным вспомогательным приспособлениям суставов относятся также внутрисуставные хрящи — диски и мениски, суставные губы, внутрикапсульные (внутрисуставные) связки. Суставы по форме подразделяются на шаровидные, эллипсовидные, цилиндрические, блоковидные, седловидные и плоские (рис. 2.11). Форма суставных поверхностей. определяет объем и направление движений, которые совершаются вокруг трех осей: фронтальной (сгибание и разгибание), сагиттальной (отведение и приведение). Вокруг вертикальной оси производится вращение, вращение внутрь называется пронацией, вращение наружу – супинацией. Если в образовании сустава участвуют две кости, — это простой сустав. Сустав, образованный тремя или несколькими костями, называется сложным. Нередко движения в двух или нескольких самостоятельных суставах происходят одновременно (правый и левый суставы нижней челюсти, суставы головки и бугорка ребра), такие суставы называются комбинированными. 13. Перечислить возбудимые ткани, их свойства. Определение понятиям «раздражимость», «возбудимость» и «возбуждение». Нервную и мышечную ткани традиционно называют возбудимыми тканями. Однако точнее будет говорить о возбудимых клетках. Например, среди структур нервной ткани возбудимостью обладают лишь нейроны. Клетки же нейроглии, которых в мозге приблизительно в 10 раз больше, чем нейронов, не обладают возбудимостью. Проводимость - способность ткани проводить возбуждение по всей своей длине. Показатель проводимости - скорость проведения возбуждения. Скорость проведения возбуждения по скелетной ткани - 6-13 м/с, по нервной ткани до 120 м/с. Проводимость зависит от интенсивности обменных процессов, от возбудимости (прямо пропорционально). Рефрактерность (невозбудимость) - способность ткани резко снижать свою возбудимость при возбуждении. В момент самой активной ответной реакции ткань становится невозбудимой. Различают: абсолютно рефрактерный период - время, в течении которого ткань не отвечает абсолютно ни на какие возбудители. Лабильность (функциональная подвижность) - способность ткани воспроизводить определенное число волн возбуждения в единицу времени в точном соответствии с ритмом наносимых раздражений. Это свойство характеризует скорость возникновения возбуждения. Показатель лабильности: максимальное количество волн возбуждения в данной ткани: нервные волокна - 500-1000 импульсов в секунду, мышечная ткань - 200-250 импульсов в секунду, синапс - 100-125 импульсов в секунду. Лабильность зависит от уровня обменных процессов в ткани, возбудимости, рефрактерности. Для мышечной ткани к четырем перечисленным свойствам добавляется пятое - сократимость. Возбудимость — свойство нервных и мышечных клеток отвечать на действие раздражителя возбуждением. Возбуждение — ответная реакция высокоспециализированных клеток на действие раздражителя, характеризующаяся проявлением специфической функции и потенциала действия этих клеток. Надо отметить, что возбудимость при эволюции высокоспециализированных клеток развилась из свойства раздражимости, является как бы частным случаем раздражимости. Раздражимость — это универсальное свойство всех клеточных структур отвечать на действие раздражителя изменением своей жизнедеятельности. Например, нейтрофильные лейкоциты, восприняв появление специфического антигена, прикрепляются к стенке капилляра и мигрируют в направлении воспалительного процесса в ткани. Эпителий кожи при воздействии ультрафиолетовых лучей изменяет обмен веществ, накапливает защитный пигмент. Возбуждение имеет специфические и неспецифические проявления. К неспецифическим проявлениям возбуждения нервных и мышечных клеток относят увеличение проницаемости клеточных мембран для минеральных ионов, ускорение обмена веществ и, соответственно, увеличение поглощения кислорода и выделения углекислого газа, снижение рН, возрастание температуры клетки и т.д. Эти проявления во многом аналогичны компонентам ответной реакции на действие раздражителя у невозбудимых клеток. Отличие лишь в том, что у возбудимых клеток эти процессы развиваются быстрее и интенсивнее. Специфическим проявлением возбуждения для мышечных клеток является сокращение, для нервных клеток — генерация и проведение потенциала действия на относительно большие расстояния, без уменьшения его амплитуды. 14. Сравнительная характеристика местного и распространяющегося возбуждения. Электротонический потенциал (местное возбуждение) • распространяется по нервным волокнам с затуханием (с декрементом ), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения; • вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см); • местное возбуждение распространяется пассивно, без затрат энергии клетки; • механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим. Потенциал действия (распространяющееся возбуждение) • распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения; • расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна; • распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов; • механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения. 15. Структура биологический мембраны, её свойства и функции. Классификация ионных каналов и их характеристика. Плазматическая мембрана, или плазмалемма, — наиболее постоянная, основная, универсальная для всех клеток мембрана. Она представляет собой тончайшую (около 10 нм) пленку, покрывающую всю клетку. Плазмалемма состоит из молекул белков и фосфолипидов (рис. 1.6). Молекулы фосфолипидов расположены в два ряда — гидрофобными концами внутрь, гидрофильными головками к внутренней и внешней водной среде. В отдельных местах бислой (двойной слой) фосфолипидов насквозь пронизан белковыми молекулами (интегральные белки). Внутри таких белковых молекул имеются каналы — поры, через которые проходят водорастворимые вещества. Другие белковые молекулы пронизывают бислой липидов наполовину с одной или с другой стороны (полуинтегральные белки). На поверхности мембран эукариотических клеток имеются периферические белки. Молекулы липидов и белков удерживаются благодаря гидрофильно-гидрофобным взаимодействиям. 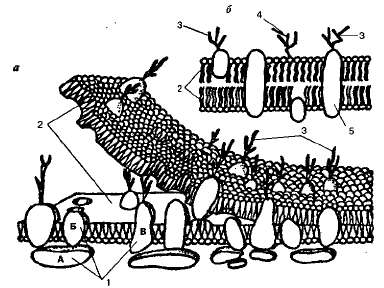 Рис.1.6. Схема строения мембраны: а — трехмерная модель; б — плоскостное изображение; 1 — белки, примыкающие к липидному слою (А), погруженные в него (Б) или пронизывающие его насквозь (В); 2 — слои молекул липидов; 3 —гликопротеины; 4 — гликолипиды; 5 — гидрофильный канал, функционирующий как пора. В состав плазматической мембраны эукариотических клеток входят также полисахариды. Их короткие, сильно развлетвленные молекулы ковалентно связаны с белками, образуя гликопротеины, или с липидами (гликолипиды). Содержание полисахаридов в мембранах составляет 2-—10% по массе. Полисахаридный слой толщиной 10—20 нм, покрывающий сверху плазмалемму животных клеток, получил название гликокаликс. Клетки с помощью распознающих участков плазмалеммы правильно ориентируются относительно друг друга, обеспечивая тем самым их сцепление и образование тканей. С распознаванием связана и регуляция транспорта молекул и ионов через мембрану, а также иммунологический ответ, в котором гликопротеины играют роль антигенов. Сахара, таким образом, могут функционировать как информационные молекулы (подобно белкам и нуклеиновым кислотам). В мембранах содержатся также специфические рецепторы, переносчики электронов, преобразователи энергии, ферментные белки. Белки участвуют в обеспечении транспорта определенных молекул внутрь клетки или из нее, осуществляют структурную связь цитоскелета с клеточными мембранами или же служат в качестве рецепторов для получения и преобразования химических сигналов из окружающей среды. Важнейшим свойством мембраны является также избирательная проницаемость. Это значит, что молекулы и ионы проходят через нее с различной скоростью, и чем больше размер молекул, тем меньше скорость прохождения их через мембрану. Это свойство определяет плазматическую мембрану как осмотический барьер. Максимальной проникающей способностью обладает вода и растворенные в ней газы; значительно медленнее проходят сквозь мембрану ионы. Диффузия воды через мембрану называется осмосом. Функции биологических мембран следующие: Отграничивают содержимое клетки от внешней среды и содержимое органелл от цитоплазмы. Обеспечивают транспорт веществ в клетку и из нее, из цитоплазмы в органеллы и наоборот. Выполняют роль рецепторов (получение и преобразование сит-налов из окружающей среды, узнавание веществ клеток и т. д.). Являются катализаторами (обеспечение примембранных химических процессов). Участвуют в преобразовании энергии Классификация ионных каналов проводится по различным параметрам и поэтому единой унифицированной классификации для них пока не существует. Так, возможна классификация по структуре (строению) и происхождению от однотипных генов. По этому принципу, например, выделяют три семейства лиганд-активируемых ионных каналов[1]: с пуриновыми рецепторами (АТФ-активируемые); с никотиновыми АХ-рецепторами, ГАМК-, глицин- и серотонин-рецепторами; с глутаматными рецепторами. При этом в одно и то же семейство попадают ионные каналы с разной ионной селективностью, а также с рецепторами к разным лигандам. Но зато образующие эти каналы белки имеют большое сходство в строении и происхождении. Ионные каналы также можно классифицировать по селективности в зависимости от проходящих через них ионов: натриевые, калиевые, кальциевые, хлорные,протонные (водородные). Согласно функциональной классификации[2], ионные каналы группируются по способам управления их состоянием на следующие виды: Неуправляемые (независимые). Потенциал-управляемые (потенциал-чувствительные, потенциал-зависимые, voltage-gated). Лиганд-управляемые (хемо-управляемые, хемочувствительные, хемозависимые, лиганд-зависимые, рецептор-активируемые). Опосредованно-управляемые (вторично-управляемые, ион-активируемые, ион-зависимые, мессенджер-управляемые, управляемые метаботропнымирецепторами). Совместно-управляемые (NMDA-рецепторно-канальный комплекс). Они открываются одновременно как лигандами, так и определённым электрическимпотенциалом мембраны. Можно сказать, что у них двойное управление. Пример: NMDA-рецепторно-канальный комплекс, имеющий сложную систему управления, включающую в себя 8 рецепторных участков-сайтов, с которыми могут связываться различные лиганды. Стимул-управляемые (механочувствительные, механосенситивные, активируемые растяжением (stretch) липидного бислоя, протон-активируемые, температурно-чувствительные). Актин-управляемые (актин-регулируемые, actin-regulated, actin-gated channels). Коннексоны (двойные поры). Наиболее часто встречаются два типа каналов: ионные каналы с лиганд-зависимыми воротами (находятся, в частности, в постсинаптической мембране нервно-мышечных соединений) и ионные каналы с потенциал-зависимыми воротами. Лиганд-зависимые каналы превращают химические сигналы, приходящие к клетке, в электрические; они необходимы, в частности, для работы химических синапсов. Потенциал-зависимые каналы нужны для распространения потенциала действия |