Метод Физиология и анатомия как науки определение, основные понятия, значение в системе медицинских знаний. Значение знаний по физиологии и анатомии человека для врачафармацевта
 Скачать 0.97 Mb. Скачать 0.97 Mb.
|

|
Неуправляемые (независимые) ионные каналы Эти каналы обычно находятся в открытом состоянии и постоянно пропускают через себя ионы за счёт диффузии по градиенту их концентрации и/или по электрическому градиенту зарядов по обе стороны мембраны. Некоторые неуправляемые каналы различают вещества и пропускают через себя по градиенту концентрации все молекулы меньше определённой величины, их называют «неселективные каналы» или «поры». Существуют также «селективные каналы», которые благодаря своему диаметру и строению внутренней поверхности переносят только определённые ионы. Примеры: калиевые каналы, участвующие в формировании мембранного потенциала покоя, хлоридные каналы, эпителиальные натриевые каналы, анионные каналы эритроцитов.[3] Потенциал-зависимые ионные каналы Эти каналы отвечают за распространение потенциала действия, они открываются и закрываются в ответ на изменение мембранного потенциала. Например, натриевые каналы. Если мембранный потенциал поддерживается на уровне потенциала покоя, натриевые каналы закрыты и натриевый ток отсутствует. Если мембранный потенциал сдвигается в положительную сторону, то натриевые каналы откроются, и в клетку начнут входить ионы натрия по градиенту концентрации. Через 0,5 мс после установления нового значения мембранного потенциала, этот натриевый ток достигнет максимума. А еще через несколько миллисекунд падает почти до 12. Во время покоя мембранного потенциала, внутриклеточная концентрация ионов натрия 12 ммоль/литр, а внеклеточная концентрация 145 ммоль/литр. Это значит, что каналы через некоторое время закрываются вследствие инактивации, даже если клеточная мембрана остается деполяризованной. Но закрывшись, они отличаются от состояния, в котором находились до открытия, теперь они не могут открываться в ответ на деполяризацию мембраны, то есть они инактивированны. В таком состоянии они останутся до тех пор, пока мембранный потенциал не вернется к исходному значению и не пройдет восстановительный период, занимающий несколько миллисекунд. Лиганд-зависимые ионные каналы Эти каналы открываются, когда медиатор, связываясь с их наружными рецепторными участками, меняет их конформацию. Открываясь, они впускают ионы, изменяя этим мембранный потенциал. Лиганд-зависимые каналы почти нечувствительны к изменению мембранного потенциала. Они генерируют электрический потенциал, сила которого зависит от количества медиатора, поступающего в синаптическую щель и времени, которое он там находится. 16.Биолектр. явления в жив.тканях; потенциал покоя. Регистрация ПП. Основн. полож. современ. теории. Происхождения биопотенциалов. ^ПП- разность потенциалов между внутренней и наружной поверхностью мембраны у клетки, которая находится в состоянии физиологического покоя. Наружная стороны заряд +, внутренняя -. Величина ПП 30-90мВ. В возникновении 3 фактора: 1. Наличие разности концентрация(градиентов) ионов K и Na между внутриклет. и внеклет. Средой. В цитоплазме концентрация иона Na в среднем 15 ммоль\л, а во внеклет. – 150ммоль\л. Ионов К внутри клетки в 30-40 раз больше, чем снаружи. Ионов Са и СI снаружи больше,чем внутри. Концентрация хлора 12-30, Са- до 10000. 2. Различная проницаемость кл. мембран для минеральных ионов.В покое проницаемость мембраны для калия больше,чем для натрия,и больше,чем для хлора.Из-за большой конц. К и высокой проницаемости для него кл.мембраны К выходит из кл. ивыносит с собой электр. Заряд +. При этом отрицательно заряж ионы не могут пройти через мембрану и ост в клетке. Отсюда, внутри заряд -, снаружи +. 3. Работа натрий-калиевого насоса.Он вносит вклад в увеличение заряда мембраны, косвенно способствует поляризации мембраны, создавая высокую конц. Калия в клетке. ^Биопотенциалы-электрические заряды, возникающие в результате жизнедеятельности клеток и тканей. Из-за биопотенциалов идет передача информации путем электрической сигнализации. Среди биопотенц.,генерируемых нервными клетками, выделяют ПП, локальный потенциал, рецепторный потенциал и ПД. Теория основывается на особенностях строения клет.биолог. мембраны, обладающей избирательной проницаемостью для ионов и других в-в из клетки во внеклет. Среду и из внеклет. В клетку. Пологается, что мембрана пронизана каналами,через которые могут проникать в клетку и из клетки ионы и др. в-в. Мембрана менее проницаема для анионов, чем для катионов, и способна изменять свою проницаемость в зависимости от ее функционального строения. 17. Биолектр. Явления в жив. Тканях: потенциал действия(ПД). График Пд и его фазы. ^ПД – быстрое изменение заряда мембраны, вызываемое действием достаточно сильных(пороговых и сверхпороговых) раздражителей. Признак наличия ПД: появление кратковременной инверсии(перемены) знака заряда на мембране.Снаружи на -0.5-2 мс становится отрицательным. ПД делится на деполяризацию (вся посходящая часть ПД, в ней выделяют участок, соответств. Локальному потенциалу, быструю деполяризацию, инверсию знака заряда), реполяризацию (ее скорость может замедляться, и этот участок называют следовой отрицательостью( или следовым отриц. Потенциалом) и гиперполризацию (возрастание поляризации мембраны – следовый полож.потенциал). 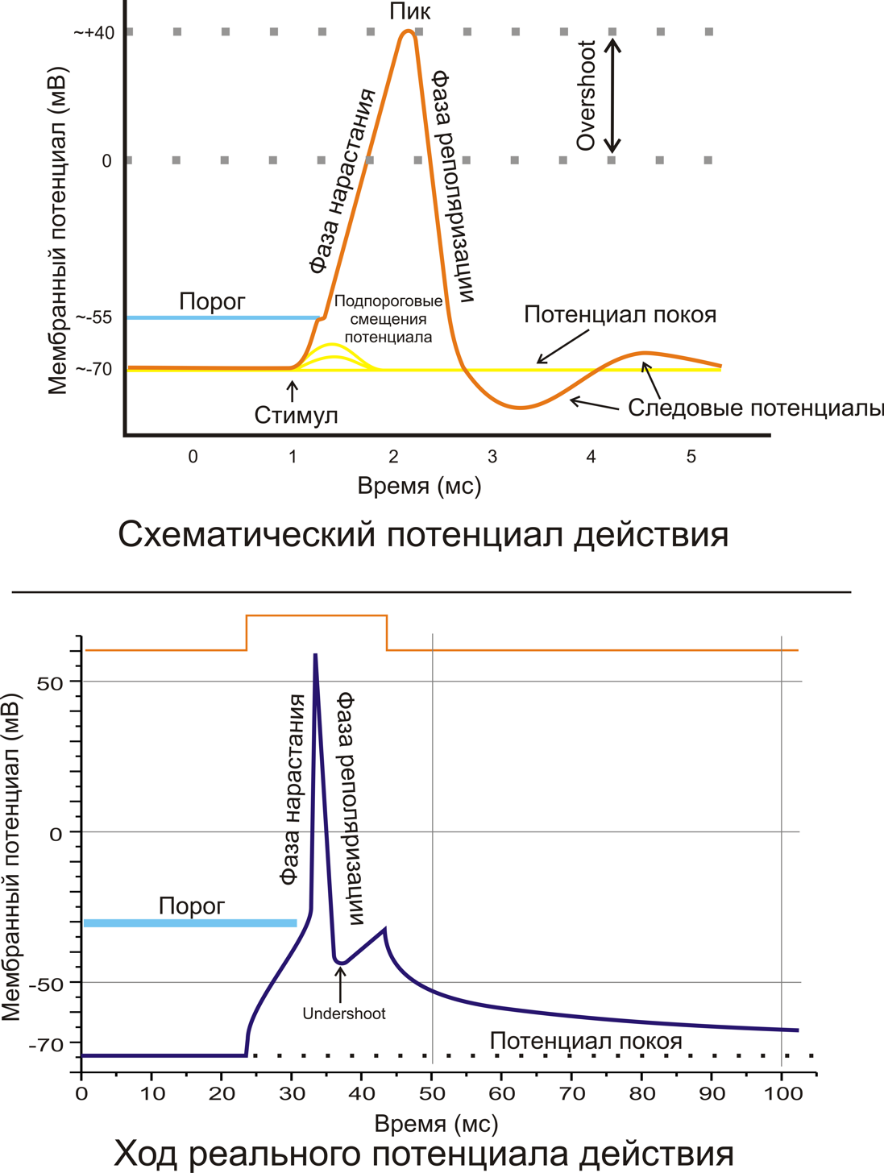 18.Изменение возбудимости мембраны клетки в различные фазы одиночного цикла возбуждения. Рефрактерность. ^Фазы одиночного цика возбуждения.1.период развития исходной деполяризации на чрезвычайно короткое время возбудимость некординально увеличивается по сопоставлению с начальной.2.развития полной деполяризации и инверсии заряда возбудимость падает до 0. Время, в течение которого отсутствует возбудимость, именуется периодом абсолютной рефрактерности. В это время даже чрезвычайно мощный раздражитель не может вызвать возбуждение ткани. 3.восстановление МП возбудимость также начинает восстанавливаться, но она еще ниже начального уровня. Время восстановления ее от 0 до начальной величины именуется периодом первичной относительной рефрактерности.Ткань может ответить возбуждением лишь на мощные, надпороговые, раздражения. 4.период экзальтации – завышенной (по сопоставлению с начальной) возбудимости. По времени он соответствует процессу реполяризации. 5.Заключительный– повторное понижение возбудимости ниже начального уровня (но не до 0), называемое периодом вторичной относительной рефрактерности.Он совпадает с развитием гиперполяризации мембраны. Возбуждение может появиться лишь в этом случае, если сила раздражения существенно превзойдет пороговую. После чего возбудимость восстанавливается, и клеточка готова к осуществлению последующего цикла возбуждения. ^Рефрактерность - период времени после возникновения на возбудимой мембране ПД, в ходе которого возбудимость мембраны снижается, а затем постепенно восстанавливается до исходного уровня. 19.Местное возбуждение (локальный ответ - ЛО), его хар-ка. Критический (пороговый) уровень деполяризации. ^ЛО –активные подпороговые изменения мембранного потенциалы, предшествующие возникновению ПД в точке раздражения нерва или мышцы.Свойства: отсутствие латентного(скрытого) периода, зависимость от силы раздражения, постепенное нарастание величины ответа и нераспространения по нервному волокну. Первые признаки проявления ЛО обнаруживаются при действии стимулов, сост 50-75% пороговой величины. Обусловлен повышением натриевой проницаемости мембраны, однако это повышение недостаточно, чтобы вызвать ПД. При пороговом стимуле ЛО перерастает в ПД. ^Пороговый уровень деполяризации - величина мембранного потенциала, при достижении которой возникает ПД. Под действием раздражения открываются «быстрые» Na каналы, вследствие чего ионы Na лавинообразно поступают в клетку. Переход положительно заряженных ионов в клетку вызывает уменьшение положительного заряда на ее наружной поверхности и увеличение его в цитоплазме. В результате этого сокращается трансмембранная разность потенциалов, а затем по мере дальнейшего поступления Na в клетку происходят перезарядка мембраны и инверсия ее заряда (поверхность становится электроотрицательной по отношению к цитоплазме) – возникает потенциал действия (ПД). Электрографическим проявлением деполяризации является спайк, или пиковый потенциал. 20. Опыты Гальвани. ^ Опыт Гальвани 1(балконный опыт). 1786г. При изучении влияния атмосферного электричества на живой организм размещал на железной решетке балкона задние лапки лягушки, закрепленные на медных крючках. При соприкосновении лапок с железной решеткой балкона наблюдалось сокращение мышц.На основании этих наблюдений Гальвани высказал мысль о существовании животного электричества. ^Опыт Гальвани 2.1794г он проделал без металла.Приподнимая нерв нервно-мышеччного препарата стеклянным крючком, он набрасывал его на поврежденный участок мышцы и наблюдал ее сокращение. Так было доказано наличие животного электричества. 21. Законы раздражения возбудимых тканей: закон силы, закон «всё или ничего». Реакция клеток, тканей на раздражитель определяется законами раздражения. Закон силы: Чем больше сила раздражителя, тем сильнее ответная реакция. Однако выраженность ответной реакции растет лишь до определенного максимума. Закону силы подчиняется целостная скелетная, гладкая мышца, так как они состоят из многочисленных мышечных клеток, имеющих различную возбудимость. Закон "всё или ничего": При подпороговых раздражениях клетки, ткани ответной реакции не возникает. При пороговой силе раздражителя развивается максимальная ответная реакция, поэтому увеличение силы раздражения выше пороговой не сопровождается ее усилением, т.е. при действии порогового и сверхпороговых раздражителей возбудимая структура отвечает одинаково. В соответствии с этим законом реагирует на раздражения одиночное нервное и мышечное волокно, сердечная мышца. 22. Законы раздражения возбудимых тканей: закон времени, хронаксия, реобаза. Закон времени действия раздражителя. При действии одной и той же силы на разные ткани потребуется разная длительность раздражения, что зависит от способности данной ткани к проявлению своей специфической деятельности, то есть возбудимости: наименьшее время потребуется для ткани с высокой возбудимостью и наибольшее время - с низкой возбудимостью. Таким образом, возбудимость обратно пропорциональна времени действия раздражителя: чем меньше время действия раздражителя, тем больше возбудимость. Порог раздражения – это минимальная сила раздражителя, при которой возникает возбуждение. Реобаза – это минимальная сила раздражителя, вызывающая возбуждение при его действии в течение неограниченно долгого времени. На практике порог и реобаза имеют одинаковый смысл. Чем ниже порог раздражения или меньше реобаза, тем выше возбудимость ткани. Полезное время – это минимальное время действия раздражителя силой в одну реобазу, за которое возникает возбуждение. Хронаксия – это минимальное время действия раздражителя силой в две реобазы, необходимое для возникновения возбуждения. Этот параметр предложил рассчитывать Л. Лапик, для более точного определения показателя времени на кривой силы-длительности. Чем короче полезное время или хронаксия, тем выше возбудимость и наоборот. 23. Законы раздражения возбудимых тканей: закон длительности, закон градиента, полярный закон. Закон длительности раздражения. Для минимального возбуждения требуется определенная критическая пороговая длительность действия раздражителя. Увеличение длительности внешнего воздействия за пределы порога ведет к нарастанию возбуждения до максимальной величины. Дальнейшее увеличение длительности действия раздражителя не ведет к нарастанию возбуждения. Эти зависимости сформулированы в законе длительности раздражения: чем длительнее раздражение, тем сильнее до известных пределов ответная реакция живой системы. Закон градиента или аккомодации. Реакция ткани на раздражение зависит от его градиента, т.е. чем быстрее нарастает сила раздражителя во времени, тем быстрее возникает ответная реакция. При низкой скорости нарастания силы раздражителя растет порог раздражения. Поэтому, если сила раздражителя возрастает очень медленно, возбуждения не будет. Это явление называется аккомодацией. Физиологическая лабильность (подвижность) – это большая или меньшая частота реакций, которыми может отвечать ткань на ритмическое раздражение. Чем быстрее восстанавливается ее возбудимость после очередного раздражения, тем выше ее лабильность. Определение лабильности предложено Н.Е. Введенским. Наибольшая лабильность у нервов, наименьшая у сердечной мышцы. Полярный закон говорит о том, что при действии на возбудимые клетки постоянного электрического тока в момент замыкания цепи возбуждение возникает в месте приложения катода, а при размыкании – в месте контакта с анодом. Воздействие на живые ткани электрическим током очень часто используется в медицинской практике. Это обусловлено тем, что пороговая сила электрического тока мала и небольшие по величине электрические потенциалы не вызывают повреждения тканей даже при многократном воздействии. В ряде случаев такой вид раздражителя близок к адекватному. Важным фактором является также то, что электрический ток легко дозировать по величине и времени. 24. Парабиоз (Н.Е.Введенский). Лабильность. Н.Е. Введенский выпустил книгу: «Возбуждение, торможение и наркоз», где показал, что живая ткань реагирует на внешние раздражители неодинаково, её поведение представляет несколько фаз. Первая фаза: «Провизорная (уравнительная) стадия» по Н.Е. Введенскому - это исчезновение различий в действии слабых и сильных ритмических раздражений. Вторая фаза: «Парадоксальная стадия» по Н.Е. Введенскому - на сильное раздражение возникает слабая реакция ткани, в ответ на слабые раздражения – более сильный ответ, чем на сильное раздражение. Третья стадия: «Тормозящая стадия» по Н.Е. Введенскому - потеря способности ткани отвечать на раздражение. Зависимость эффектов раздражения нерва от силы тока обусловлена тем, что при повышении силы стимулов увеличивается число возбужденных нервных волокон и возрастает частота импульсов, возникающих в каждом волокне, так как сильный стимул способен вызвать залп импульсов. Таким образом, нерв реагирует высокой частотой возбуждений в ответ на сильное раздражение. При развитии парабиоза способность к воспроизведению частых ритмов, т. е. лабильность, падает. При малой силе или редком ритме раздражений каждый импульс, возникший в неповрежденном участке нерва, проводится и через парабиотический участок, так как к моменту его прихода в эту область возбудимость, сниженная после предыдущего импульса, полностью успевает восстановиться. При сильном раздражении, когда импульсы следуют друг за другом с большой частотой, каждый следующий импульс, приходящий в парабиотический участок, попадает в стадию относительной рефрактерности после предыдущего. В эту стадию возбудимость волокна резко снижена, а амплитуда ответа уменьшена. Поэтому распространяющееся возбуждение не возникает, а происходит лишь еще большее снижение возбудимости. В участке парабиоза быстро приходящие друг за другом импульсы как сами себе преграждают путь. В уравнительную фазу парабиоза все эти явления выражены ещё слабо, поэтому происходит лишь трансформация частого ритма в более редкий. В результате эффекты частых (сильных) и относительно редких (умеренных) по силе раздражений уравниваются, парадоксальную же стадию циклы восстановления возбудимости настолько затягиваются, что частые (сильные) раздражения вообще оказываются малоэффективными. С особой наглядностью эти явления могут быть прослежены на одиночных нервных волокнах при раздражении их стимулами разной частоты. Так, И.Тасаки воздействовал на один из перехватов Ранвье миелинового нервного волокна лягушки раствором уретана и исследовал проведение через такой перехват нервных импульсов. Он показал, что, в то время как редкие стимулы проходили через перехват беспрепятственно, частые задерживались им. Н. Е. Введенский рассматривал парабиоз как особое состояние стойкого, неколеблющегося возбуждения, как бы застывшего в одном участке нервного волокна. Он полагал, что волны возбуждения, приходящие в этот участок из нормальных частей нерва, как бы суммируются с имеющимися здесь «стационарным» возбуждением и углубляют его. Такое явление Н. Е. Введенский рассматривал как прообраз перехода возбуждения в торможение в нервных центрах. Торможение, по мысли Н. Е. Введенского,— это результат «перевозбуждения» нервного волокна или нервной клетки. Лабильность – функциональная подвижность возбудимой ткани. Мерой лабильности является максимальное число волн возбуждения, которые может генерировать ткань в единицу времени. Величина лабильности зависит от длительности протекания одиночной волны возбуждения и длительности фазы абсолютной рефрактерности. Так, вставочные нейроны спинного мозга могут воспроизводить более 500 волн возбуждения в секунду. У них высокая лабильность. А мотонейроны, передающие импульсацию к мышцам, способны генерировать не более 100 волн возбуждения в секунду – у них более низкая лабильность. 25. Рецепция: виды и свойства, общие механизмы действия. Биологическое значение рецепции. Рецепция - восприятие и преобразование энергии раздражителей в нервное возбуждение, осуществляемое рецепторами. Клеточная и сенсорная рецепция Под клеточной рецепцией понимают процесс восприятия и преобразования химического сигнала в сложную последовательность внутриклеточных химических процессов. Клеточная рецепция обеспечивает возможность обмена информации между клетками, который осуществляется при помощи биологически активных веществ (гормонов, медиаторов). Обязательным этапом такого межклеточного взаимодействия является связывание молекул вещества с соответствующей молекулой клетки-мишени, называемой клеточным рецептором . Роль клеточных рецепторов играют специфические белковые молекулы, которые могут быть расположены на поверхности клетки, в цитоплазме или в ядре. Сенсорной рецепцией называют процесс восприятия и преобразования энергии раздражителей внешней и внутренней среды организма в энергию нервных импульсов, передаваемую по чувствительным нервам в ЦНС. Сенсорный рецептор представляет собой нервную клетку или комплекс нервной и эпителиальной клетки, специально приспособленный для восприятия определенного типа раздражителей. Сенсорные рецепторы являются начальными звеньями любой рефлекторной дуги, а также участвуют в оценке параметров полезного приспособительного результата в функциональных системах организма. |