|
|
Сборник всех лекций. Общая характеристика обмена веществ и энергии. Биоэнергетика
План:
1. Распад сложных углеводов пищи и гликогена.
2. Распад глюкозы
2.1. Дихотомический путь.
2.2. Апотомический путь.
3. Глюконеогенез и гликогенез.
(Сл.) Схема превращений углеводов
1. Распад сложных углеводов пищи и гликогена
Человек и животные, в отличие от растений, не способны к биосинтезу углеводов из неорганических веществ и получают углеводы из пищи.
Гидролиз сложных углеводов пищи начинаетсяпод действием ферментов слюны и поджелудочной железы (амилазы и мальтазы) и завершается в тонком кишечнике.
В слюне присутствует гидролитический фермент α-амилаза (α-1,4-гликозидаза), расщепляющая в крахмале α-1,4-гликозидные связи. В ротовой полости не может происходить полное расщепление крахмала, так как действие фермента на крахмал кратковременно. Кроме того, амилаза слюны не расщепляет α-1,6-гликозидные связи (связи в местах разветвлений), поэтому крахмал переваривается лишь частично с образованием крупных фрагментов - декстринов и небольшого количества мальтозы. Следует отметить, что амилаза слюны не гидролизует гликозидные связи в дисахаридах.
Действие амилазы слюны прекращается в резко кислой среде содержимого желудка (рН 1,5-2,5). Желудочный сок не содержит ферментов, расщепляющих углеводы. В желудочном содержимом возможен лишь незначительный кислотный гидролиз гликозидных связей.
α-Амилаза поджелудочной железы, так же, как α-амилаза слюны, действует как эндогликозидаза. Панкреатическая α-амилаза не расщепляет α-1,6-гликозидные связи в крахмале. Гидролиз 1,6-связей в точках ветвления цепей амилопектина осуществляет амило-1,6-глюкозидаза.
Амилазы не расщепляют гликозидные связи, которыми соединены остатки глюкозы в молекуле целлюлозы. Целлюлоза проходит через кишечник непереваренной, в толстом кишечнике она может подвергаться действию бактериальных ферментов и частично расщепляться с образованием спиртов, органических кислот и СО2.
Мальтоза, изомальтоза и триозосахариды, образующиеся в верхних отделах кишечника из крахмала, - промежуточные продукты, а также дисахариды пищи сахароза и лактоза гидролизуются под действием специфических дисахаридаз в тонком кишечнике (мальтазы, изомальтазы, инвертазы (сахаразы) и лактазы).
Совместное действие всех перечисленных ферментов завершает переваривание пищевых олиго- и полисахаридов с образованием моносахаридов, основной из которых - глюкоза.
Схема гидролиза крахмала:
крахмал →декстрины (полисахариды с меньшим числом структурным звеньев) → мальтоза → α-D-глюкоза.
Кроме глюкозы, из углеводов пищи также образуются фруктоза и галактоза, в меньшем количестве - манноза, ксилоза, арабиноза.
Моносахариды всасываются в тонком кишечнике в кровь и доставляются воротной веной в печень, где часть глюкозы откладывается в виде гликогена. Часть глюкозы обеспечивает энергией реакции, происходящие в печени. Кроме того, определенное количество глюкозы превращается в жиры.
Регуляция уровня глюкозы в крови осуществляется ЦНС и эндокринной системой. Он сохраняется постоянным в пределах 3,3-5,8 ммоль/л крови.
Наряду с гидролизом сложных углеводов, поступающих с пищей, важную роль в поддержании нормального уровня глюкозы играет распад гликогена. Гликоген – основной запасающий углевод у животных и человека.
Гликогенолиз – процесс анаэробного распада гликогена до глюкозы –осуществляется под действием фермента в печени.
Фосфоролиз гликогена происходит с образованием глюкозо-1-фосфата (основной путь) (фермент гликогенфосфорилаза в печени и мышцах).
В мышцах гликоген расщепляется быстро, когда появляется потребность в АТФ при мышечной работе. В печени гликоген расщепляется медленнее, когда в крови снижается уровень глюкозы и надо его поднять до нормального.
2. Распад глюкозы
2.1. Дихотомический путь
Дихотомический путь заключается в распаде глюкозы на две трёхуглеродные молекулы. Гликолиз – ферментативный процесс, в ходе которого глюкоза превращается в пировиноградную кислоту (пируват или ПВК) с одновременным образованием АТФ.
В анаэробных (в отсутствии кислорода) пируват переходит в молочную кислоту.
В аэробных условиях (в присутствии кислорода) пируват включается в цикл трикарбоновых кислот и окисляется в митохондриях до углекислого газа и воды.
Значение гликолиза:
- в анаэробных условиях гликолиз – единственный процесс в организме животных, растений и многих микроорганизмов, приводящий к образованию АТФ;
- в аэробных условиях гликолиз составляет начальную стадию деструкции углеводов, связанной далее с циклом трикарбоновых кислот.
Стадии гликолиза
1. Подготовительная стадия
В гликолиз вовлекаются различные гексозы, в основном глюкоза; молекулы других гексоз превращаются в глюкозо-6-фосфат (Г-6-Ф), который изомеризуется во фруктозо-6-фосфат(Ф-6-Ф). Первая стадия заканчивается образованием глицеральдегид-3-фосфата (ГАФ).
1. Фосфорилирование глюкозы – пусковая стадия гликолиза (фермент гексокиназа):

АТФ + глюкоза → АДФ + глюкоза-6-фосфат (Г-6-Ф)
2. Изомеризация глюкозо-6-фосфата во фруктозо-6-фосфат (Ф-6-Ф) (гексозофосфатизомераза):

Г-6-Ф Ф-6-Ф
3. Фосфорилирование фруктозо-6-фосфата во фруктозо-1,6-дифосфат (фермент 6-фосфофруктокиназа); это наиболее медленная реакция гликолиза, определяющая его скорость:

Ф-6-Ф + АТФ → Ф-1,6-ДФ + АДФ
4. Фруктозо-1,6-дифосфат расщепляется на две фосфотриозы (фермент альдолаза):
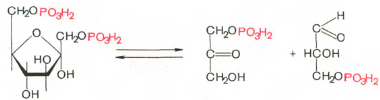
Ф-1,6-ДФ Дигидрокси- Глицеральдегид-
ацетонфосфат 3-фосфат
(ДГАФ) (ГАФ)
5. Изомеризация триозофосфатов (фермент триозофосфатизомераза):

ДГАФ ГАФ
В реакции гликолиза включается только глицеральдегидфосфат. На первой стадии гликолиза молекула глюкозы путем двух фосфорилирований и расщепления превратилась в две молекулы фосфотриоз.
2. Окислительная стадия гликолиза (генерация АТФ по механизму субстратного фосфорилирования)
Вторая стадия гликолиза включает окислительно-восстановительные реакции и реакции фосфорилирования, в результате которых регенерируется АТФ. Глицеральдегид-3-фосфат окисляется до пировиноградной кислоты или молочной кислоты с образованием АТФ и НАДН.
6. Реакция образования 1,3-дифосфоглицериновой кислоты
(1,3-ДФГК), представляющего собой высокоэнергетическое соединение:
Н-С=О глицеральдегидфос- О=С-О-РО3Н2
| фодегидрогеназа |
Н-С-ОН + Н3РО4+ НАД+ ----------------- Н-С-ОН + НАДН
| |
СН2ОРО3Н2 СН2О-РО3Н2
ГАФ 1,3-ДФГК
7. Образование АТФ и 3-фосфоглицериновой кислоты (3-ФГК):
О=С-ОРО3Н2 НО-С=О
| фосфоглицераткиназа |
Н-С-ОН + АДФ → Н-С-ОН + АТФ
| |
СН2О-РО3Н2 СН2О-Р О3Н2
1,3-ДФГК 3-ФГК
В отличие от окислительного фосфорилирования образование АТФ из высокоэнергетических соединений называется субстратным фосфорилированием.
8. Внутримолекулярный перенос фосфатной группы:
НО-С=О НО-С=О
| фосфоглицеромутаза |
Н-С-ОН → Н-С-О-РО3Н2
| |
СН2О-РО3Н2 СН2ОН
3-ФГК 2-ФГК
9. 2-фосфоглицериновая кислота в результате отщепления молекулы воды переходит в фосфоенолпировиноградную кислоту (фосфоенолпируват), а фосфатная связь в положении 2 становится высокоэргической (фермент енолаза):

2-ФГК фосфоенолпируват
10. Разрыв высокоэргической связи и перенос фосфатного остатка от фосфоенолпирувата на АДФ (субстратное фосфорилирование) катализируется ферментом пируваткиназой:

фосфоенолпируват ПВК
11. В анаэробных условиях происходит восстановление пировиноградной кислоты и образуется лактат (при участии фермента лактатдегидрогеназы и кофермента НАДН):

ПВК Лактат
Молочная кислота в анаэробных условиях является конечным продуктом гликолиза, например в мышцах. В печени 1/5 часть молочной кислоты окисляется до СО2 и Н2О в аэробной фазе дыхания и 4/5 регенерируется в гликоген.
Суммарное уравнение гликолиза в анаэробных условиях:
С6Н12О6 + 2АДФ + 2Н3РО4 → 2СН3СН(ОН)СООН + 2АТФ + 2Н2О
Суммарное уравнение гликолиза в аэробных условиях:
С6Н12О6+2АДФ+2Н3РО4+2НАД+→ 2СН3СОСООН+2АТФ+2Н2О+2НАДН∙Н+
В реакциях гликолиза при превращениях 1 моль глюкозы запасается 2 моль АТФ. Его к.п.д.- 35%.
Брожение
Многие микроорганизмы (бактерии, дрожжевые и плесневые грибы) получают энергию за счет брожения. Брожение является наиболее примитивным способом получения энергии.
Разнообразные брожения совпадают с гликолизом во многих стадиях.
При молочнокислом брожении водород переносится на пируват с образованием лактата:
лактатдегидрогеназа
СН3-СО-СООН + НАДН + Н+ → СН3-СН(ОН)-СООН3 + НАД+
При спиртовом брожении пируват вначале декарбоксилируется с образованием уксусного альдегида и углекислого газа под действием фермента пируватдекарбоксилазы (необходимы ионы магния и тиаминпирофосфат):
СН3-СО-СООН → СО2 + СН3-СНО
Уксусный альдегид превращается в спирт под действием алкогольдегидрогеназы:
СН3-СНО + НАДН + Н+ → СН3-СН2ОН + НАД+
Брожение может идти в анаэробных и аэробных условиях. В присутствии кислорода может идти окисление спирта до уксусной кислоты. Также известны маслянокислое, пропионовокислое, муравьинокислое и лимоннокислое брожение. В организме человека процессы брожения наблюдаются в кишечнике.
2.2. Апотомический путь распада глюкозы
Ключевую роль в данном процессе играют пентозофосфаты, поэтому он называется также пентозофосфатным циклом. Он выполняет важные метаболические функции: служит источником НАДФН, необходимых для реакций восстановления, и пентоз – предшественников нуклеотидов, а также в аэробных условиях может быть источником энергии.
Пентозофосфатный путь имеет две фазы.
Первая фаза - окислительная:
Д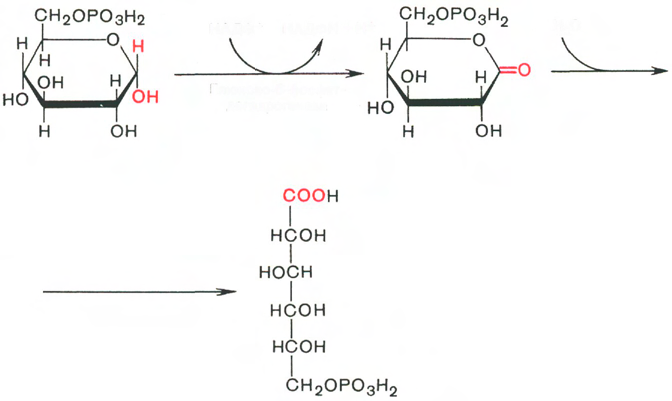 егидрирование глюкозо-6-фосфата (фермент глюкозо-6-фосфатдегидрогеназа) и гидролиз образовавшегося 6-фосфоглюконолактона до 6-фосфоглюконовойкислоты (6-ФГК): егидрирование глюкозо-6-фосфата (фермент глюкозо-6-фосфатдегидрогеназа) и гидролиз образовавшегося 6-фосфоглюконолактона до 6-фосфоглюконовойкислоты (6-ФГК):
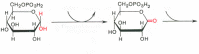
Г-6-Ф 6-фосфоглюконолактон 6-ФГК
Дегидрирование и декарбоксилирование 6-фосфоглюконовой кислоты приводит к образованию рибулезо-5-фосфата:
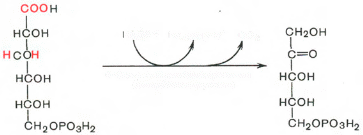
6-ФГК рибулезо-5-Ф
Таким образом, глюкозо-6-фосфат последовательно окисляется до пентозофосфатов.
Вторая фаза - неокислительная. Она представляет собой взаимопревращения сахарофосфатов, в результате которых регенерируется глюкозо-6-фосфат.
Посредством пентозофосфатного цикла может происходить полное окисление глюкозо-6-фосфата до углекислого газа. При этом шесть молекул глюкозо-6-фосфата дают пять молекул глюкозо-6-фосфата и шесть молекул углекислого газа.
Суммарное уравнение пентозофосфатного цикла:
6Г-6-Ф + 7Н2О + 12НАДФ → 5Г-6-Ф + 6СО2 + 12НАДФН2 + Н3РО4
или:
Г-6-Ф + 7Н2О + 12НАДФ → 6СО2 + 12НАДФН2 + Н3РО4
Синтезируется 12·3 = 36 молекул АТФ на 1 молекулу глюкозы.
Такое полное окисление глюкозо-6-фосфата происходит не во всех клетках. Чаще всего на одном из этапов пентозофосфатный путь переходит в гликолитический. Существует три пути сопряжения с гликолизом на уровне глюкозо-6-фосфата, фруктозо-6-фосфата и глицеральдегид-3-фосфата.
3. Глюконеогенез и гликогенез
Глюконеогенез и гликогенез относятся к основным анаболическим процессам углеводного обмена.
Глюконеогенез – процесс синтеза глюкозы из неуглеводных предшественников. Процесс протекает в печени, менее интенсивно в корковом слое почек и слизистом эпителии кишечника.
Главная функция глюконеогенеза заключается в поддержании уровня глюкозы в крови во время голодания и интенсивной физической работы. Постоянное поступление глюкозы в качестве источника энергии особенно необходимо для нервной ткани и эритроцитов. Недостаток глюкозы в крови прежде всего ощущает головной мозг, который не может обеспечить потребность в энергии за счет метаболизма других энергоёмких веществ.
Субстратами глюконеогенеза являются пируват, лактат, глицерол, аминокислоты (схема 1). Включение различных субстратов в глюконеогенез зависит от физиологического состояния организма:
- лактат является продуктом анаэробного гликолиза в эритроцитах, работающих мышцах и других тканях с низким содержанием О2;
- глицерол высвобождается при гидролизе жиров в жировой ткани в постабсорбтивный период или при физической нагрузке;
- аминокислоты образуются в результате распада белков мышц и соединительной ткани и включаются в глюконеогенез при длительном голодании или продолжительной мышечной нагрузке. Аминокислоты, участвующие в глюконеогенезе, называются гликогенными; к ним относят аминокислоты, при метаболических превращениях которых образуется пируват, оксалоацетат, фосфоенолпируват; это практически все аминокислоты, кроме лейцина.
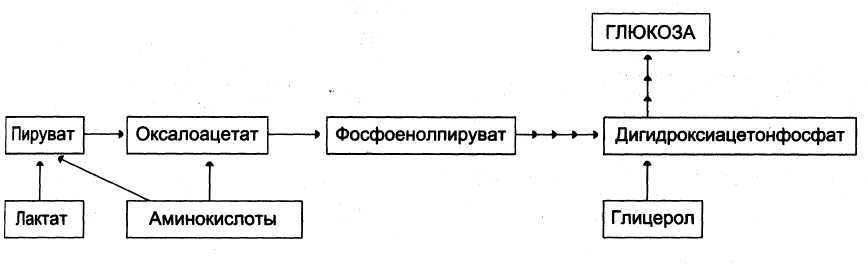
Схема 1. Предшественники для глюконеогенеза
Большинство реакций глюконеогенеза протекает за счет обратимых реакций гликолиза и катализируется теми же ферментами.
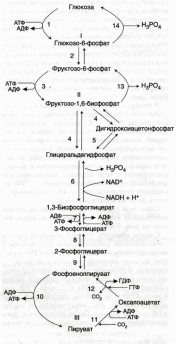
Схема гликолиза (слева) и глюконеогенеза (справа)
Обходные реакции глюконеогенеза
Три реакции гликолиза необратимы вследствие происходящих в ходе их значительных энергетических сдвигов:
• фосфорилирование глюкозы;
• фосфорилирование фруктозо-6-фосфата;
• превращение фосфоенолпирувата в пируват.
Обход этих энергетических барьеров обеспечивают ключевые ферменты глюконеогенеза, не участвующие в гликолизе. Четыре необратимые реакции глюконеогенеза называют обходными реакциями глюконеогенеза:
Первые две необратимые реакции глюконеогенеза - фосфорилирование пирувата с образованием промежуточного соединения - оксалоацетата, который затем превращается в фосфоенолпируват. Катализатором превращения пирувата в оксалоацетат является митохондриальный фермент – пируваткарбоксилаза (коферментом пируваткарбоксилазы является биотин (витамин Н)). Реакция образования оксалоацетата протекает в митохондриях, роль её заключается также в пополнении фонда оксалоацетата для цикла Кребса.
Все последующие реакции глюконеогенеза протекают в цитоплазме. Мембрана митохондрий непроницаема для оксалоацетата, и он переносится в цитоплазму в виде других метаболитов: малата или аспартата. В цитоплазме указанные соединения вновь переходят в оксалоацетат. Затем оксалоацетат декарбоксилируется и фосфорилируется под действием фосфоенолпируваткарбоксикиназы и образуется фосфоенолпируват.
Суммарное уравнение фосфорилирования пирувата:
Пируват + АТФ + ГТФ → Фосфоенолпируват + АДФ + ГДФ + Н3РО4
К необратимым реакциям глюконеогенеза относятся также реакции дефосфорилирования фруктозо-1,6-дифосфата (он превращается во фруктозо-6-фосфат) и дефосфорилирования глюкозо-6-фосфата (он, в свою очередь, превращается в глюкозу). Отщепление остатков фосфорной кислоты происходит под действием соответствующих фосфатаз.
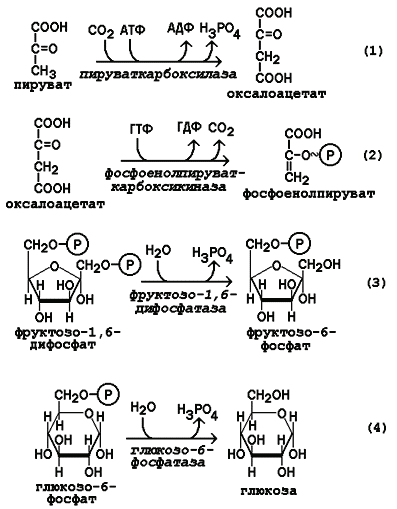
В ходе синтеза 1 моль глюкозы из 2 моль пирувата расходуется 4 моль АТФ и 2 моль ГТФ. Суммарное уравнение
2Пируват + 4АТФ + 2ГТФ +2НАДН∙Н+ +4Н2О → Глюкоза + 2НАД+ +
+ 4АДФ +2 ГДФ + 6Н3РО4
Гликогенез (биосинтез гликогена) начинается с затраты молекулы АТФ для фосфорилирования глюкозы с образованием глюкозо-6-фосфата и её изомеризации с образованием глюкозо-1-фосфата (Г-1-Ф).
При взаимодействии (Г-1-Ф) и уридинтрифосфата (УТФ) образуется уридинфосфатглюкоза (УДФ-глюкоза), которая и служит источником остатков глюкозы при гликогенезе. Фермент гликогенсинтетаза сн имает остаток глюкозы с УДФ-глюкозы и переносит его на конец растущей цепи гликогена.
Гликоген синтезируется главным образом в печени и составляет до 6 % от массы печени, в скелетных мышцах - 0,5-2 %. Масса мышц значительно больше массы печени, поэтому в скелетных мышцах сосредоточено около 2/3 от всего гликогена тела.
|
|
|
 Скачать 3.68 Mb.
Скачать 3.68 Mb.