biokhimia_Vosstanovlen (Восстановлен). Предмет и задачи биологической химии. Обмен веществ и энергии, сложная структурная организация, гомеостаз и самовоспроизведение как важнейшие признаки живой материи
 Скачать 2.96 Mb. Скачать 2.96 Mb.
|

Методы определение глюкозы в кровиОпределение глюкозы в крови – важный этап, чтобы диагностировать сахарный диабет. Для этого необходимо определить состояние углеводного обмена, а в первую очередь содержание глюкозы в крови. В норме ее концентрация лежит в пределах 3, 3-5, 5 ммоль/л. Существует огромное количество методик определения, позволяющих установить глюкозы в крови. Среди них можно выделить редуктометрические, колориметрические, ферментативные методы определения:
63. Аэробный гликолиз. Последовательность реакций до образования пирувата (аэробный гликолиз). Физиологическое значение аэробного гликолиза. Использование глюкозы для синтеза жиров. Катаболизм глюкозы - основной поставщик энергии для процессов жизнедеятельности организма. А. Основные пути катаболизма глюкозы Окисление глюкозы до СО2 и Н2О (аэробный распад). Аэробный распад глюкозы можно выразить суммарным уравнением: С6Н12О6 + 6 О2 → 6 СО2 + Н2О + 2820 кДж/моль. Этот процесс включает несколько стадий (рис. 7-33).
Б. Аэробный гликолиз Аэробным гликолизом называют процесс окисления глюкозы до пировиноградной кислоты, протекающий в присутствии кислорода. Все ферменты, катализирующие реакции этого процесса, локализованы в цитозоле клетки. 1. Этапы аэробного гликолиза В аэробном гликолизе можно выделить 2 этапа.
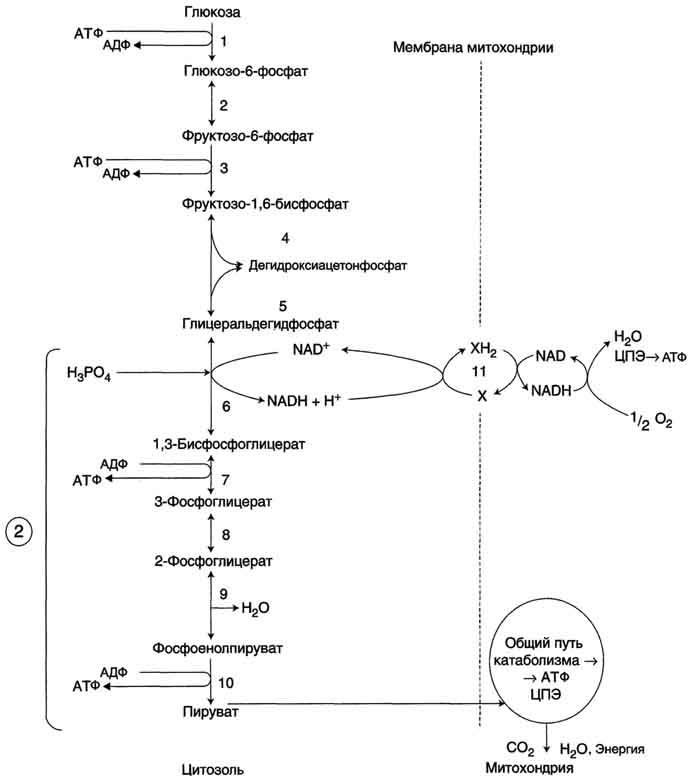 Аэробный распад глюкозы. 1-10- реакции аэробного гликолиза; 11 - малат-аспартатный челночный механизм транспорта водорода в митохондрии; 2 (в кружке) - стехиометрический коэффициент.  общий путь катаболизма; 4 - аэробный распад глюкозы; 5 - анаэробный распад глюкозы (в рамке); 2 (в кружке) - стехиометрический коэффициент." align=bottom width=357 height=179 border=0> общий путь катаболизма; 4 - аэробный распад глюкозы; 5 - анаэробный распад глюкозы (в рамке); 2 (в кружке) - стехиометрический коэффициент." align=bottom width=357 height=179 border=0>Пути катаболизма глюкозы. 1 - аэробный гликолиз; 2, 3 - общий путь катаболизма; 4 - аэробный распад глюкозы; 5 - анаэробный распад глюкозы (в рамке); 2 (в кружке) - стехиометрический коэффициент. 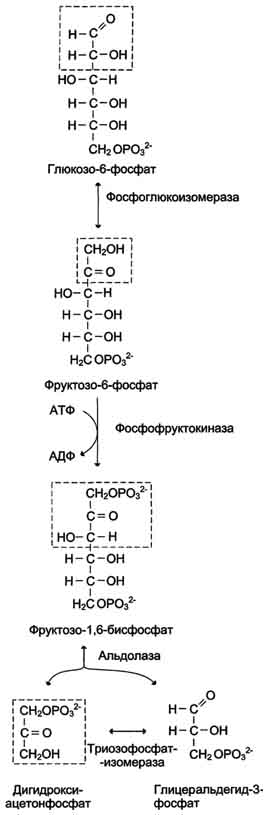 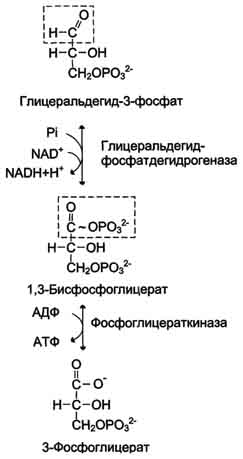 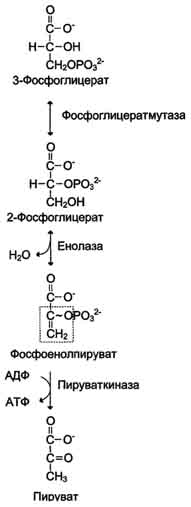 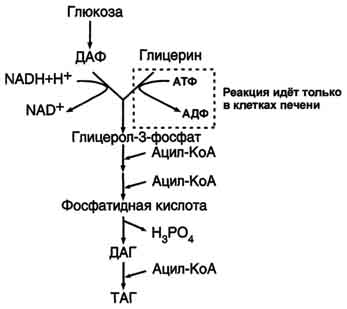 64. Анаэробный гликолиз. Реакция гликолитической оксидоредукции; субстратное фосфорилирование. Распространение и физиологическое значение анаэробного распада глюкозы. Анаэробным гликолизом называют процесс расщепления глюкозы с образованием в качестве конечного продукта лактата. Этот процесс протекает без использования кислорода и поэтому не зависит от работы митохондриальной дыхательной цепи. АТФ образуется за счёт реакций субстратного фосфорилирования. Суммарное уравнение процесса: С6Н1206 + 2 Н3Р04 + 2 АДФ = 2 С3Н6О3 + 2 АТФ + 2 Н2O. 1. Реакции анаэробного гликолиза При анаэробном гликолизе в цитозоле протекают все 10 реакций, идентичных аэробному гликолизу. Лишь 11-я реакция, где происходит восстановление пирувата цитозольным NADH, является специфической для анаэробного гликолиза. Восстановление пирувата в лактат катализирует лактатдегидро-геназа (реакция обратимая, и фермент назван по обратной реакции). С помощью этой реакции обеспечивается регенерация NAD+ из NADH без участия митохондриальной дыхательной цепи в ситуациях, связанных с недостаточным снабжением клеток кислородом. Роль акцептора водорода от NADH (подобно кислороду в дыхательной цепи) выполняет пируват. Таким образом, значение реакции восстановления пирувата заключается не в образовании лактата, а в том, что данная цитозольная реакция обеспечивает регенерацию NAD+. К тому же лактат не является конечным продуктом метаболизма, удаляемым из организма. Это вещество выводится в кровь и утилизируется, превращаясь в печени в глюкозу, или при доступности кислорода превращается в пируват, который вступает в общий путь катаболизма, окисляясь до СО2 и Н2О. Строение лактатдегидрогеназы, механизм действия и значение определения активности этого фермента для диагностики заболеваний описывались ранее в разделе 2. Баланс АТФ при анаэробном гликолизе Анаэробный гликолиз по сравнению с аэробным менее эффективен. В этом процессе катаболизм 1 моль глюкозы без участия митохондриальной дыхательной цепи сопровождается синтезом 2 моль АТФ и 2 моль лактата. АТФ образуется за счёт 2 реакций субстратного 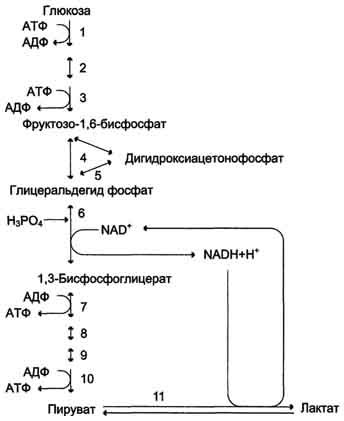 Рис. 7-40. Анаэробный гликолиз. фосфорилирования. Поскольку глюкоза распадается на 2 фосфотриозы, то с учётом стехиометри-ческого коэффициента, равного 2, количество моль синтезированного АТФ равно 4. Учитывая 2 моль АТФ, использованных на первом этапе гликолиза, получаем конечный энергетический эффект процесса, равный 2 моль АТФ. Таким образом, 10 цитозольных ферментов, катализирующих превращение глюкозы в пируват, вместе с лактатдегидрогеназой обеспечивают в анаэробном гликолизе синтез 2 моль АТФ (на 1 моль глюкозы) без участия кислорода. Анаэробный распад глюкозы происходит в мышцах, в первые минуты мышечной работы, в эритроцитах (в которых отсутствуют митохондрии), а также в разных органах в условиях ограниченного снабжении их кислородом, в том числе в клетках опухолей. Для метаболизма клеток опухолей характерно ускорение как аэробного, так и анаэробного гликолиза. Но преимущественный анаэробный гликолиз и увеличение синтеза лактата служит показателем повышенной скорости деления клеток при недостаточной обеспеченности их системой кровеносных сосудов. 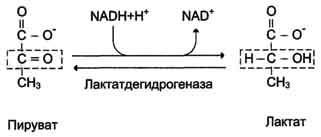 65. Биосинтез глюкозы (глюконеогенез) из аминокислот, глицерина и молочной кислоты; регуляция глюконеогенеза. Биотин, роль в образовании оксалоацетата. Взаимосвязь гликолиза в мышцах и глюконеогенеза в печени (цикл Кори). 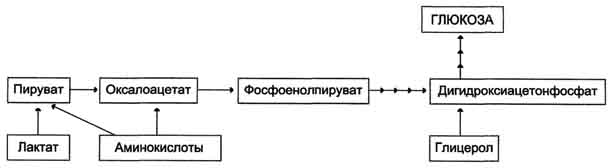 Рис. 7-44. Включение субстратов в глюконеогенез. 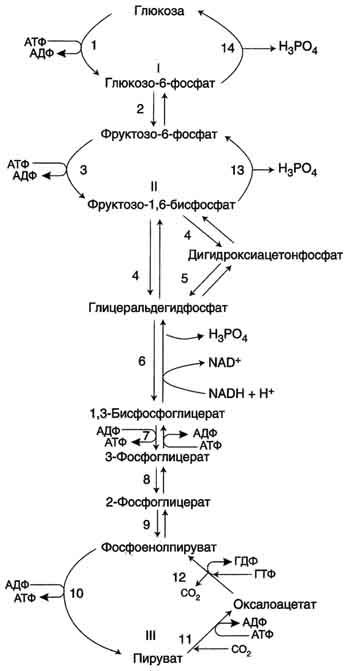 Рис. 7-45. Гликолиз и глюконеогенез. Ферменты обратимых реакций гликолиза и глюконеогенеза: 2 - фосфоглюкоизоме-раза; 4 - альдолаза; 5 - триозофосфатизомераза; 6 - глицеральдегидфосфатдегидрогеназа; 7 -фосфоглицераткиназа; 8 - фосфоглицератмутаза; 9 - енолаза. Ферменты необратимых реакций глюконеогенеза: 11 - пируваткарбоксилаза; 12 - фосфоенолпируваткарбоксикиназа; 13 - фруктозо-1,6-бисфосфатаза; 14 -глюкозо-6-фосфатаза. I-III -субстратные циклы. 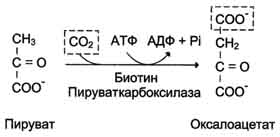 Рис. 7-46. Образование оксалоацетата из пирувата. 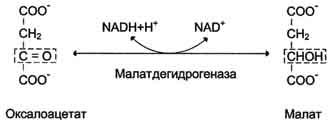 Рис. 7-47. Превращение оксалоацетата в малат. 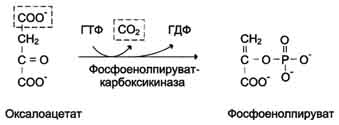 Рис. 7-48. Превращение оксалоацетата в фосфоенолпируват. 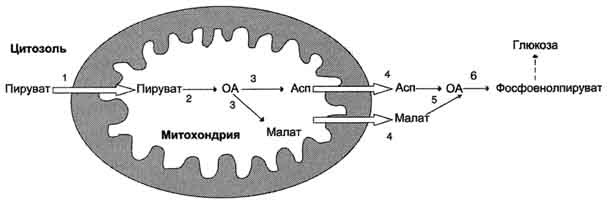 Рис. 7-49. Образование оксалоацетата, транспорт в цитозоль и превращение в фосфоенолпируват. 1 - транспорт пирувата из цитозоля в митохондрию; 2 - превращение пирувата в оксалоацетат (ОА); 3 - превращение ОА в малат или аспартат; 4 - транспорт аспартата и малата из митохондрии в цитозоль; 5 - превращение аспартата и малата в ОА; 6 - превращение ОА в фосфоенолпируват.  печени с током крови в работающую мышцу; 4 - использование глюкозы как энергетического субстрата сокращающейся мышцей и образование лактата." align=bottom width=477 height=180 border=0> печени с током крови в работающую мышцу; 4 - использование глюкозы как энергетического субстрата сокращающейся мышцей и образование лактата." align=bottom width=477 height=180 border=0>Рис. 7-50. Цикл Кори (глюкозолактатный цикл). 1 - поступление лактата из сокращающейся мышцы с током крови в печень; 2 - синтез глюкозы из лактата в печени; 3 - поступление глюкозы из печени с током крови в работающую мышцу; 4 - использование глюкозы как энергетического субстрата сокращающейся мышцей и образование лактата. 66. Гликоген, биологическое значение. Биосинтез и мобилизация гликогена. Регуляция синтеза и распада гликогена. Многие ткани синтезируют в качестве резервной формы глюкозы гликоген. Синтез и распад гликогена обеспечивают постоянство концентрации глюкозы в крови и создают депо для её использования тканями по мере необходимости. Гликоген - разветвлённый гомополимер глюкозы, в котором остатки глюкозы соединены в линейных участках α-1,4-гликозидной связью. В точках ветвления мономеры соединены α-1,6-гликозидными связями. Эти связи образуются примерно с каждым десятым остатком глюкозы. Следовательно, точки ветвления в гликогене встречаются примерно через каждые десять остатков глюкозы. Так возникает древообразная структура с молекулярной массой >107Д, что соответствует приблизительно 50 000 остатков глюкозы (рис. 7-21). Таким образом, в молекуле гликогена имеется только одна свободная аномерная ОН-группа и, следовательно, только один восстанавливающий (редуцирующий) конец. В клетках животных гликоген - основной резервный полисахарид. При полимеризации глюкозы снижается растворимость образующейся молекулы гликогена и, следовательно, её влияние на осмотическое давление в клетке. Это обстоятельство объясняет, почему в клетке депонируется гликоген, а не свободная глюкоза. Гликоген хранится в цитозоле клетки в форме гранул диаметром 10-40 нм. С гранулами связаны и некоторые ферменты, участвующие в метаболизме гликогена, что облегчает их взаимодействие с субстратом. Разветвлённая структура гликогена обусловливает большое количество концевых мономеров, что способствует работе ферментов, отщепляющих или присоединяющих мономеры при распаде или синтезе гликогена, так как эти ферменты могут одновременно работать на нескольких ветвях молекулы. Гликоген депонируется главным образом в печени и скелетных мышцах. После приёма пищи, богатой углеводами, запас гликогена в печени может составлять примерно 5% от её массы. В мышцах запасается около 1% гликогена, однако масса мышечной ткани значительно больше и поэтому общее количество гликогена в мышцах в 2 раза больше, чем в печени. Гликоген может синтезироваться во многих клетках, например в нейронах, макрофагах, клетках жировой ткани, но содержание его в этих тканях незначительно. В организме может содержаться до 450 г гликогена. Распад гликогена печени служит в основном для поддержания уровня глюкозы в крови в постабсорбтивном периоде. Поэтому содержание гликогена в печени изменяется в зависимости от ритма питания. При длительном голодании оно снижается почти до нуля. Гликоген мышц служит резервом глюкозы - источника энергии при мышечном сокращении. Мышечный гликоген не используется для поддержания уровня глюкозы в крови. Как уже упоминалось ранее, в клетках мышц нет фермента глюкозо-6-фосфатазы, и образование свободной глюкозы невозможно. Расход гликогена в мышцах зависит в основном от физической нагрузки . 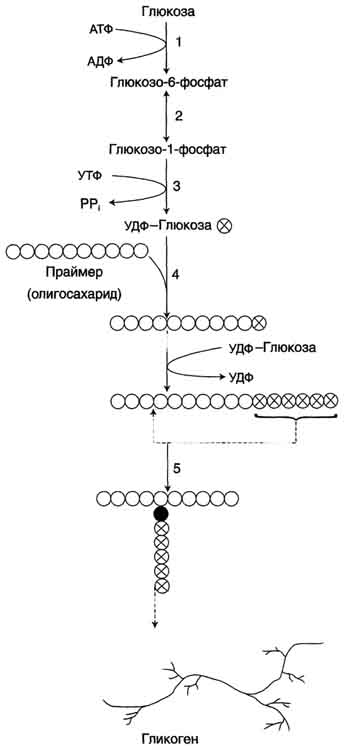 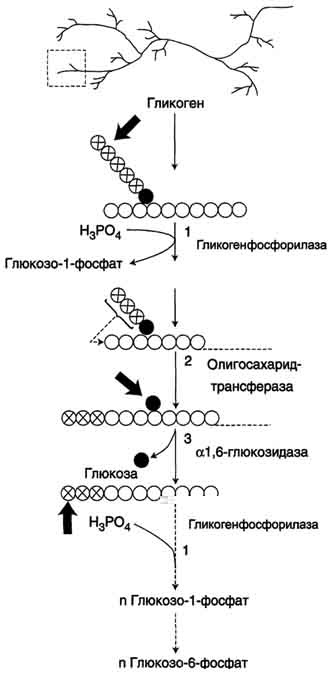 Рис. 7-23. Синтез гликогена. 1 - глюкокиназа или гексокиназа; 2 - фосфоглюкомутаза; 3 - УДФ-глюкрпирофосфорилаза; 4 - гликогенсинтаза (глюкозилтрансфераза); 5 - фермент "ветвления" (амило-1,4 → 1,6-глюкозилтрансфераза), светлые и заштрихованные кружки - глюкозные остатки, закрашенные кружки - глюкозные остатки в точке ветвления. Рис. 7-25. Распад гликогена. В рамке - фрагмент гликогена с точкой ветвления. Закрашенный кружок - глюкозный остаток, связанный α-1,6-гликозидной связью; светлые и заштрихованные кружки - глюкозные остатки в линейных участках и боковых ветвях, связанные α-1,4-гликозидной связью. 1 - Гликогенфосфорилаза; 2 - олигосахаридтрансфераза; 3 - α-1,6-глюкозидаза 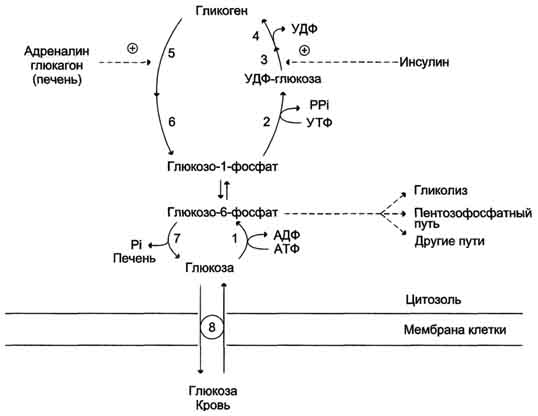 |