10.2.Экспрессия трансгенов
Если трансгены в своем функционировании проявляют тканеспецифичность, то уровень их экспрессии зависит от места интеграции в хромосому. В тех редких случаях, когда экспрессия трансгена полностью отсутствует, это объясняют его интеграцией в гетерохроматиновые участки хромосом, неактивные в отношении транскрипции, или другими эффектами положения. Поскольку уровень экспрессии трансгенов у некоторых трансгенных мышей может превышать таковой у его эндогенного гомолога, делают вывод об отсутствии необходимости в точной локализации гена на хромосоме для его эффективной экспрессии. Тканеспецифический характер экспрессии генов обеспечивают, главным образом, их энхансеры – цис-действующие регуляторные последовательности нуклеотидов, которые располагаются как внутри, так и вне генов на значительном удалении от них (см. рис. I.30). Обнаружены гены, экспрессирующиеся в клетках только одного типа (строгая тканеспецифическая экспрессия), в клетках немногих тканей и в клетках многих или большинства типов (так называемые гены "домашнего хозяйства"). В том случае, если энхансеры обеспечивают абсолютный тканеспецифический характер экспрессии, контекст окружающих трансген последовательностей нуклеотидов хромосомы оказывает влияние на уровень его экспрессии только в клетках одного типа. Если же энхансеры дают возможность функционировать трансгену в клетках разных тканей, то уровень экспрессии будет варьировать в клетках этих тканей даже при интеграции в геном единственной его копии.
Обнаружена слабая корреляция между числом копий трансгенов, тандемно интегрированных в геном животного, и суммарным уровнем их экспрессии. Это указывает на функциональную активность лишь немногих трансгенов из всего кластера их множественных копий.
Таким образом, данные, полученные к настоящему времени, указывают на то, что экспрессия рекомбинантных генов у трансгенных животных в большой степени напоминает таковую, происходящую в природных условиях в гомологичном генетическом окружении. Объединение структурных частей рекомбинантных генов с конкретными регуляторными последовательностями организма-реципиента позволяет получать строго детерминированную, тканеспецифическую экспрессию этих генов в процессе трансгеноза.
10.3.Использование трансгенов у животных
Техника трансгеноза открывает практически безграничные, принципиально новые возможности исследования экспрессии генов. Ниже будут кратко рассмотрены четыре активно развиваемые направления использования трансгенов в фундаментальных и прикладных исследованиях. К ним относятся: а) изучение молекулярных механизмов экспрессии генов; б) исследование механизмов дифференцировки соматических клеток в онтогенезе многоклеточных организмов; в) подходы к изменению физиологического статуса лабораторных и сельскохозяйственных животных; г) моделирование наследственных и приобретенных заболеваний человека. Два последних направления исследований являются фундаментом для современной и будущей генотерапии.
10.3.1.Исследование механизмов экспрессии генов
Как уже упоминалось выше, цис-действующие регуляторные последовательности нуклеотидов обеспечивают тканеспецифический характер экспрессии трансгенов. Этим свойством воспользовались для определения точной локализации таких регуляторных элементов генов. Например, с использованием делеционного картирования был идентифицирован энхансерный элемент гена эластазы-1 крыс. Полный ген эластазы-1 в составе фрагмента ДНК длиной в 7,2 т.п.о. экспрессировался у трансгенных мышей, что указывало на наличие в данном фрагменте ДНК всех последовательностей нуклеотидов, необходимых для функционирования гена. Удаление с помощью делеций 5’-фланкирующих последовательностей нуклеотидов этого гена не влияло на тканеспецифический характер его экспрессии в ацинарных клетках поджелудочной железы мышей (место обычной активации гена) до тех пор, пока не затрагивалась последовательность длиной в 205 п.о., непосредственно прилегающая к 5’-концевой его части (рядом с точкой инициации транскрипции). Делеции, которые оставляли лишь 72 п.о. 5’-фланкирующей последовательности, полностью инактивировали ген, что указывало на расположение регуляторного элемента между нуклеотидами в положениях -205 и -72. Такая последовательность обладала всеми свойствами энхансера и сохраняла активность после перемещения за 3 т.п.о. перед сайтом ее нормальной локализации или встраивания в интрон. При этом для нее была характерна способность активировать гетерологичные промоторы.
Использование таких эффективных, хотя и трудоемких методов иногда является единственным путем идентификации тканеспецифических регуляторных элементов генов. Дело в том, что клетки в культуре тканей часто претерпевают дедифференцировку и утрачивают изначально присущий им тканеспецифический характер экспрессии своих генов, поэтому применение культур тканей вместо трансгенных животных для таких целей проблематично.
Экспрессия трансгенов обнаруживается по появлению соответствующего им белкового продукта или транскрипта, а также по специфическим физиологическим реакциям организмов-реципиентов. Это позволяет использовать трансгеноз для изучения путей и механизмов дифференцировки соматических клеток в биологии развития, а также для изменения физиологической и физической конституции сельскохозяйственных животных, моделирования многих наследственных заболеваний человека и разработки методов их лечения.
10.3.2.Токсигены в исследовании дифференцировки соматических клеток в онтогенезе
Выше рассматривалась возможность применения гибридных токсинов для специфического воздействия на группы соматических клеток, обладающих определенными фенотипическими маркерами. Высокоспецифической элиминации определенных групп дифференцирующихся клеток можно достигнуть с использованием трансгеноза. В этом случае гены каталитических субъединиц токсинов (например A-субъединиц дифтерийного токсина (DT-A) или рицина) с помощью генно-инженерных приемов помещают под контроль регуляторных генетических элементов, обеспечивающих их тканеспецифическую экспрессию в клетках трансгенных экспериментальных животных.
После интеграции таких токсигенов в геном развивающегося эмбриона они передаются от клетки к клетке в рядах клеточных поколений без экспрессии до тех пор, пока клетки не начинают дифференцироваться с образованием регуляторных белков, трансактивирующих регуляторные последовательности токсигенов. С этого момента токсигены активированы, т.е. приобрели способность направлять биосинтез белков-токсинов, которые избирательно поражают дифференцирующиеся клетки. Иными словами, начав дифференцироваться в определенном направлении, соматические клетки совершают самоубийство. Поскольку каталитические субъединицы обладают токсическим эффектом лишь внутри соматических клеток и сами по себе не могут проникнуть в окружающие их клетки извне, их токсический эффект распространяется только на клетки, в которых происходит экспрессия токсигенов.
С помощью такой стратегии были получены впечатляющие результаты при изучении путей терминальной дифференцировки соматических клеток животных in vivo. В частности, были продемонстрированы различия во временнóй активации промоторов генов 2- и 2-кристаллинов в процессе формирования хрусталиков глаз у мышей, а также последствия активации промоторов гена эластазы-I, специфически функционирующего в ацинарных клетках поджелудочной железы.
Из-за большой токсичности вышеупомянутых белковых токсинов (чтобы убить клетку, достаточно одной молекулы) использование таких токсигенов может сопровождаться артефактами и приводить к ранней гибели трансгенных эмбрионов. Для преодоления этих трудностей были разработаны методы, при которых сам по себе трансген не оказывает токсического действия на клетки, но внутриклеточное присутствие белкового продукта его экспрессии делает клетки чувствительными к определенному экзогенному воздействию.
В одной из таких систем использовали ген тимидинкиназы вируса простого герпеса (HSV-tk), продукт которого, в отличие от тимидинкиназы клеток-хозяев, обладает способностью фосфорилировать некоторые аналоги нуклеозидов, например ацикловир и FIAU [1-(2-дезокси-2-фтор--D-арабинофуранозил)-5-иодоурацил]. После фосфорилирования с помощью HSV-tk до нуклеозидмонофосфатов образующиеся аналоги нуклеотидов могут далее фосфорилироваться тимидилаткиназой клеток-хозяев с образованием соответствующих дезоксирибонуклеозидтрифосфатов и конкурентно блокировать репликацию ДНК соматических клеток. Аналогичный подход был использован для элиминации в эмбриогенезе клеток, экспрессирующих кристаллины, кроветворных клеток (в частности тимоцитов) и в ряде других случаев.
Токсигены находят применение не только для изучения генетики развития, но и для создания животных, моделирующих наследственные и приобретенные заболевания человека.
10.3.3.Изменение физиологического статуса лабораторных и сельскохозяйственных животных
Одними из первых указаний на возможность использования трансгеноза для изменения физиологических параметров и физической конституции организма животных были результаты работ по экспрессии трансгенов гормонов, контролирующих рост млекопитающих: рилизинг-фактора гормона роста (GRF), соматостатина и самого гормона роста (growth hormone, GH). Гормон роста индуцирует образование в печени инсулиноподобного фактора роста I (IGF-I), который, в свою очередь, ускоряет пролиферацию клеток. Повышенное внутриклеточное содержание GH крысы, быка или человека в клетках трансгенных мышей сопровождалось ускорением их роста через три недели после рождения, рост стабилизировался через 12 недель, когда размер мышей в два раза превышал обычный. К аналогичному эффекту приводила экспрессия у трансгенных мышей гена GRF человека, находящегося под контролем промотора гена металлотионеина, который стимулировал эндогенное образование GH, что сопровождалось повышением внутриклеточного уровня IGF-I-мРНК и ускорением роста трансгенных животных.
Увеличение уровня экспрессии GH у трансгенных мышей обусловливает и ряд других физиологических реакций организма. Например, под действием трансгена GH отмечены ингибирование биосинтеза эндогенного GH и дегенерация соматотропных клеток гипофиза, в норме продуцирующих этот гормон. Экспрессия приводила, кроме того, к изменению половых различий в строении печени, ослаблению фертильности самок и преждевременному старению трансгенных животных.
Результаты таких опытов легли в основу целого направления прикладных исследований, в которых были предприняты попытки изменения физиологического статуса сельскохозяйственных животных путем гормональных воздействий, опосредованных трансгенами. Были получены трансгенные овцы и свиньи, однако результаты оказались не вполне удовлетворительными из-за вышеупомянутых побочных эффектов GH. Решение этой проблемы, по-видимому, лежит на пути создания трансгенных животных с регулируемой экспрессией трансгенов. М. Мак-Грейну с соавторами недавно удалось получить гигантских мышей с помощью трансгена GH быков, экспрессирующегося под контролем регуляторной области крысиного гена З-энолпируваткарбоксикиназы, для которого характерна тканеспецифическая экспрессия в печени и почках. Авторам удалось управлять экспрессией трансгена путем изменения содержания углеводов и белков в пищевом рационе мышей. Такой же подход был успешно применен для получения полноценных трансгенных свиней, которые характеризовались ускоренным увеличением веса и существенным уменьшением содержания жира в тканях.
Другое направление биотехнологии, в котором используется трансгеноз, – изучение возможности биосинтеза чужеродных белков в клетках молочных желез млекопитающих с последующей их секрецией в молоко. Идея этого подхода заключается в применении молочных желез крупных сельскохозяйственных животных в качестве биореакторов для биосинтеза биологически активных пептидов и белков. Предполагалось, что в результате экспрессии трансгенов в клетках молочных желез удастся синтезировать рекомбинантные белки, претерпевшие правильные посттрансляционные модификации (в частности гликозилирование), что, как правило, не удается делать в других генетических системах.
Уже в первых опытах с трансгенами тканевого активатора плазминогена (tPA) человека, интегрированными в геном в виде кДНК, была показана возможность специфической экспрессии его в молочных железах мышей, причем содержание функционально активного tPA составляло около 0,1 мг на 1 мл. Похожие результаты были получены с трансгенами фактора IХ коагуляции крови, 1-антитрипсина, -интерферона, фолликулостимулирующего гормона и интерлейкина 2 человека, а также ряда других белков, имеющих значение для биотехнологии.
Во всех вышеперечисленных направлениях исследований усилия были сосредоточены на получении трансгенных животных-биореакторов, секретирующих в молоко белковые продукты экспрессии трансгенов, что предполагает их дальнейшую очистку. В отличие от этого в других исследованиях предпринимались попытки изменения формулы и питательных свойств молока. В частности, Дж. Симмонсу с соавторами удалось получить экспрессию гена -лактальбумина овец в молочных железах трансгенных мышей. Авторы не исключали принципиальной возможности сделать состав молока крупных сельскохозяйственных животных более пригодным для новорожденных детей или людей, обладающих повышенной чувствительностью к лактозе.
10.3.4.Моделирование наследственных и приобретенных заболеваний человека
Для разработки эффективных методов лечения наследственных и приобретенных заболеваний человека, а также для полного понимания их этиологии требуется моделирование соответствующих симптомов на лабораторных животных. В этом случае проблема заключается в направленном введении мутаций в гены организма животных, инактивация которых приводит к развитию патологических процессов. Разработка эффективных методов получения трансгенных животных позволила исследователям вплотную подойти к решению данного вопроса.
Основой для решения послужили две работы, выполненные М. Хупером и М. Куэном с соавторами и опубликованные в 1987 г. Авторам удалось разработать общий подход к избирательной инактивации генов в организме животных. Мишенью для инактивации стал ген гипоксантин-гуанозин-фосфорибозилтрансферазы (ГГФРТ), нефункциональное состояние которого у человека приводит к развитию заболевания Леша–Нихана. Мышей, дефектных по гену ГГФРТ, получали из эмбриональных стволовых клеток линии ES, инактивируя ген с помощью спонтанных мутаций или интегрируя в него геном ретровирусов (разновидность инсерционного мутагенеза). Клетки с инактивированным геном ГГФРТ удобно отделять от клеток дикого типа на селективной питательной среде в присутствии 6-тиогуанина. Позднее было показано, что инактивация гена ГГФРТ может быть достигнута путем гомологичной рекомбинации гена дикого типа с мутантным геном или его частью, которые вводят в клетки ES с помощью электропорации или микроинъекций в составе линеаризованных векторных плазмид. Основная проблема, возникающая при инактивации гена-мишени с помощью гомологичной рекомбинации, заключается в низкой частоте (
10-6) рекомбинационных событий, приводящих к правильной интеграции экзогенной последовательности нуклеотидов в инактивируемый ген. Эта проблема в настоящее время решается путем отбора требуемых мутантных клеток из общей популяции клеток, в которых интеграция инактивирующего вектора в хромосомы произошла случайным образом.
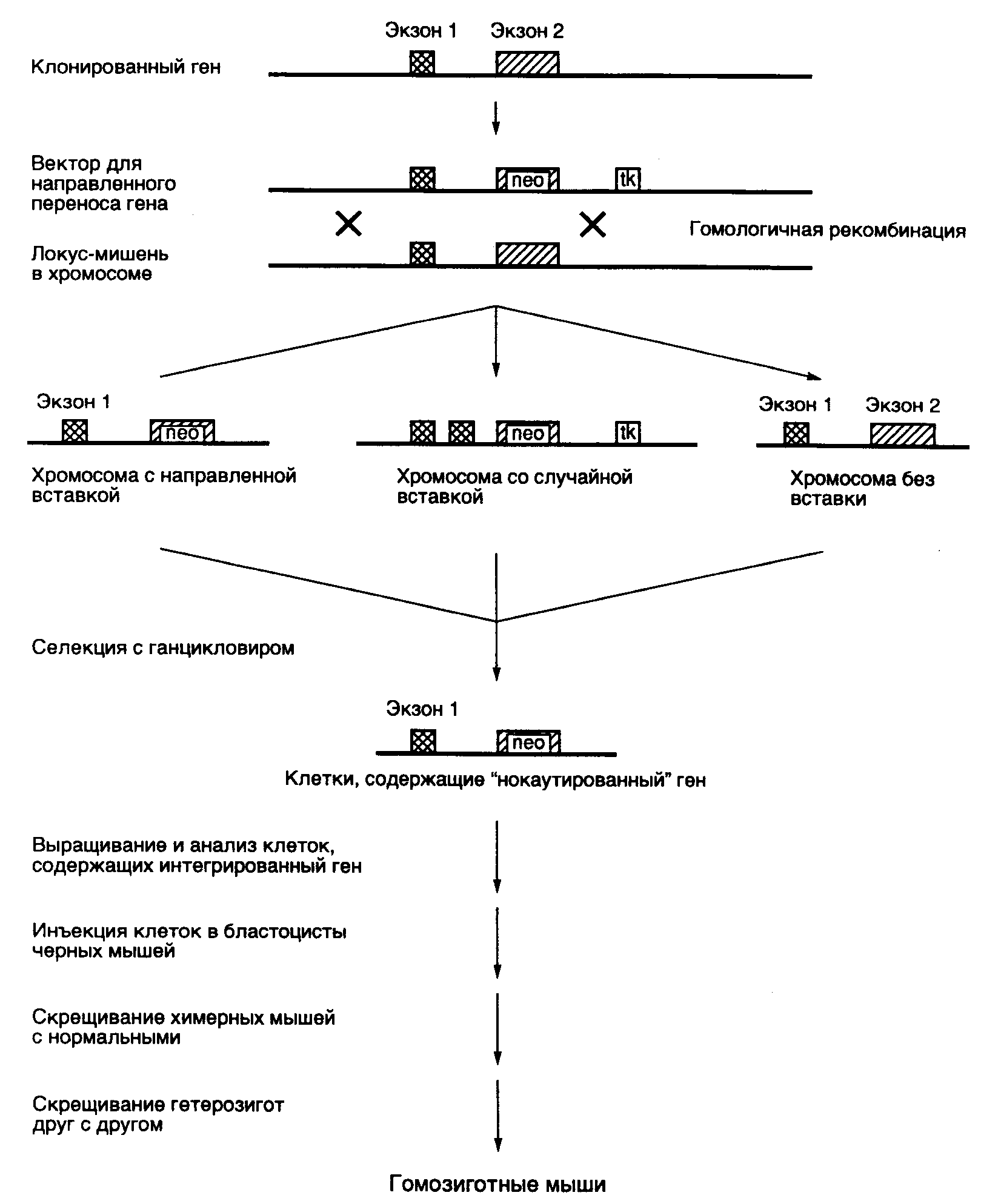
Рис. II.29. Схема адресной доставки генов в геном эмбриональных стволовых клеток с использованием гомологичной рекомбинации ("генный нокаут")
Показано положение гена устойчивости к неомицину (neo), интегрированного в экзон 2 инактивируемого гена, который используется для позитивного отбора, а также гена тимидинкиназы вируса простого герпеса (tk), используемого для негативной селекции клеток в присутствии ганцикловира
Чтобы создать селективные условия отбора, в последовательности нуклеотидов рекомбинантной ДНК, гомологичные последовательностям инактивируемого гена, вводят доминантный селектируемый маркер в виде гена устойчивости к антибиотикам (например neo), который экспрессируется только в случае правильной интеграции в ген-мишень под контроль его регуляторных элементов. Другая, более эффективная система отбора основана на одновременном проведении негативной и позитивной селекции клеток, у которых произошла правильная интеграция инактивирующего вектора. В этой системе помимо доминантного селектируемого маркера neo инактивирующий вектор содержит другой маркерный ген, экспрессия которого приводит к гибели клеток ES на селективной среде. Маркерным геном часто является ген тимидинкиназы вируса простого герпеса (HSV-tk), принцип действия которого был рассмотрен выше. Ген HSV-tk вводится в инактивирующий вектор таким образом, что он удаляется в результате гомологичной рекомбинации и сохраняется в хромосоме в случае неспецифической интеграции рекомбинантной ДНК, что приводит к цитотоксическому эффекту. Обобщенная схема направленного введения мутаций в гены животных in vivo, которые приводят к их инактивации во всех клетках трансгенного организма ("генный нокаут"), представлена на рис. II.29.
Разработка универсальных методов направленной инактивации любого требуемого гена во всех клетках организма трансгенных животных позволила за короткое время создать модели таких наследственных заболеваний человека, как -талассемия, мышечная дистрофия Дюшенна, серповидно-клеточная анемия, муковисцидоз, болезнь Леша–Нихана и целого ряда других распространенных синдромов. Создание этих методов открыло также возможности генотерапии наследственных заболеваний путем замещения в клетках зародышевой линии мутантных аллелей на нормальные. Результаты, полученные в этой области, будут обсуждаться в разделе 10.5.
Следует подчеркнуть, что инактивация генов-мишеней у лабораторных животных – не единственный способ моделирования наследственных и особенно приобретенных заболеваний человека, поскольку причиной многих заболеваний является не отсутствие функционирования, а сверхэкспрессия определенных генов. Например, высокий уровень экспрессии трансгена аполипопротеина AII сопровождается развитием острого атеросклероза у трансгенных мышей. Только исчерпывающее знание этиологии заболевания допускает его адекватное моделирование, и, наоборот, лишь создание адекватных моделей может убедить исследователя в том, что он правильно понимает его природу.
|
 Скачать 5.83 Mb.
Скачать 5.83 Mb.