В.А. Догель - Зоология беспозвоночных. Тип губки (spongia, или porifera)
 Скачать 15 Mb. Скачать 15 Mb.
|

|
Палеонтология. Наиболее древние брюхоногие, известные с кембрия, - относятся к переднежаберным. Среди них интересны еще сохранившие 470 симметричную раковину Bellerophontidae (ордовик — пермь) и дожившая с триаса до наших дней Pleurotomaria (рис. 462,Л). В карбоне появляются заднежаберные и легочные моллюски. Такие распространенные сейчас пресноводные роды, как прудовики и катушки, ведут свое начало с юры (p. Lymnaea) и мела (p. Planorbis). Классификация. Большое количество видов брюхоногих моллюсков распределяется между тремя подклассами. Классификация в пределах последних отличается сложностью в связи с большим разнообразием в строении брюхоногих. ПОДКЛАСС I. ПЕРЕДНЕЖАБЕРНЫЕ (PROSOBRANCHIA) Имеется хиастоневрия. Внутренностный мешок испытал поворот на 180° или близкий к этому. Жабра одна (реже две), лежит впереди сердца. Преимущественно раздельнополы. Встречаются в море, реже в пресных водах или на суше. На ноге у многих форм имеется крышечка. Включает два отряда. Отряд I. Древние брюхоногие, или Двупредсердные (Archaeogastropoda, или Dioto- cardia). Самые примитивные брюхоногие моллюски, обладающие двумя предсердиями и педальными нервными стволами: образование педальных ганглиев еще не произошло. Мантийный комплекс симметричен только у самых примитивных форм (подотр. Zygo-branchia), имеющих два ктенидия, два предсердия и две почки. Такова, например, Pleurotomaria, обитающая в Индийском океане, турбоспиральная раковина ее несет длинную вырезку, начинающуюся от устья (рис. 462, Л). Это самая древняя форма из современных брюхоногих. У морского уха — Haliotis (рис. 462, В) — становится заметной асимметрия мантийного комплекса: правый ктенидий меньше левого. Haliotis, обитающий в тропических морях, обладает характерной уплощенной уховидной раковиной, пронизанной рядом небольших отверстий. Еще более просто устроены колпачковчд-ные раковины некоторых форм (например, Fissurella; рис. 462, Б),имеющие вид небольшого конуса с отверстием на вершине. Намечающаяся в пределах этой группы асимметрия мантийного комплекса находит свое окончательное выражение в подотр. Azygo-branchia, представители которого полностью лишены правого ктенидия. Это преимущественно морские тропические формы, как, например, обладающий высокой турбоспи-ральной раковиной Trochus — обитатель прибрежной зоны, часто встречающийся на коралловых рифах. Имеются формы, вторично перешедшие к жизни в пресных водах, — в ряде рек СССР обычен Theodoxus fluviatilis. У форм подотряда Docoglossa, обладающих небольшой колпачковидной раковиной, также сохраняется один ктенидий, часто замещающийся вторичными жабрами. Типичный представитель этой группы — широко распространенный обитатель приливно-отливной зоны Patella (рис. 462, Г). Отряд 2. Однопредсердные (Monotocardia). Мантийный комплекс представителей этой группы слагается только из органов левой стороны — один ктенидий, предсердие и почка. У ряда форм и, в первую очередь, перешедших к жизни в пресной воде или на земле ктенидий либо исчезает совсем, либо замещается вторичными жабрами. Имеются педальные ганглии. В настоящее время отр. Monotocardia многими исследователями разбивается на два самостоятельных отряда: значительная часть более примитивных форм относится к отр. Mesogastropoda, в то время как высшие представители передне-жаберных составляют отр. Stenoglossa. К отряду Однопредсердных относятся: Littorina — многочисленные виды этого рода обитают на литорали, многие легко переносят пребывание на воздухе во время отлива; Сургаеа — тропические морские улитки с красивой инволютной раковиной; Мигех (рис. 462, Д),выделяющий слизистый секрет (в древности применялся для окраски тканей в пурпурный цвет); Conus (рис. 462, Е) с ядовитыми железами и зубами радулы. В северной части Атлантического и Тихого океанов распространен Buccinum — один из обычных съедобных переднежаберных моллюсков. В озерах и прудах часто встречаются пресноводные переднежаберные — Viviparus и Bithynia; от пресноводных легочных моллюсков эти формы легко отличаются по присутствию на ноге крышечки. Своеобразно измененными морскими однопредсердными переднежаберными являются киленогие (Heteropoda), ведущие планктонный образ жизни; средняя часть ноги сплющена с боков и образует выдающийся вниз плавник, задняя часть ноги вытягивается в виде хвоста. Киленогие встречаются преимущественно в теплых морях, хищники. К ним относятся: Carinaria с маленькой колпачковидной раковиной; Pterotrachea — без раковины (см. рис. 430). Среди однопредсердных имеется несколько форм, паразитирующих на иглокожих, например Parenteroxenos dogieli — паразит голотурии Сиси-maria, достигающий в длину более 1 м. 471 ПОДКЛАСС II. ЗАДНЕЖАБЕРНЫЕ (OPISTHOBRANCHIA) Нервная система эпиневральная. Мантийный комплекс испытал сдвиг из переднего положения на правую сторону тела. Обычно имеется один ктенидий, лежащий позади сердца. Предсердие одно. Гермафродиты. Раковина часто редуцирована. Исключительно морские формы. Включают 2 отряда: покрытожаберных (Tectibranchia) и голожаберных (Nudibran-chia). У покрытожаберных ктенидий развит и обычно имеется раковина. В наших северных морях распространены Scaphander, Cylichna — формы с инволютной раковиной. К этой же группе относятся и крылоногие моллюски (Pteropoda), нога которых изменена в два боковых плавника. В северных морях обычен морской ангел — Clione limacina (см. рис. 440) — без раковины, оранжево-красного цвета, служит пищей беззубым китам. У представителей отряда голожаберных ктенидий заменен вторичными жабрами. Раковина отсутствует. По внешнему виду они часто билатерально симметричны. Представители: Dendronotus, Catriona (см. рис. 443) —с многочисленными жабрами на спине, широко распространены в наших северных морях. ПОДКЛАСС III. ЛЕГОЧНЫЕ (PULMONATA) Нервная система эутиневральная. Жабры, как правило, заменены легким. Гермафродиты. Все органы мантийного комплекса непарны. Наземные и пресноводные формы. Крышечки нет. Подкласс включает два , отряда: сидячеглазых (Basommatophora) и стебельчатоглазых (Stylom-matophora). Первый отряд представлен главным образом пресноводными формами, глаза располагаются у основания второй пары щупалец. К ним относится большинство наших пресноводных брюхоногих: обыкновенный прудовик — Lymnaea stagnalis (см. рис. 438), малый прудовик — Lym-паеа truncatula — промежуточный хозяин печеночной двуустки (с. 170), роговая катушка — Planorbarius corneas и т. д. Отряд стебельчатоглазых объединяет преимущественно наземные формы, у которых глаза помещаются на вершине второй пары щупалец. Представители: янтарка — Succinea — обитает в сырых местах на траве и кустарниках, промежуточный хозяин сосальщика Leucochloridium (см. рис. 145). Широко распространена виноградная улитка (Helix pomatia). Ощутимый вред сельскому хозяйству могут наносить слизни Deroceras, Parmacella и др. КЛАСС III. ПЛАСТИНЧАТОЖАБЕРНЫЕ, ИЛИ ДВУСТВОРЧАТЫЕ (LAMELLIBRANCHIA, ИЛИ BIVALVIA) Пластинчатожаберные образуют большой класс (около 20000 видов) морских и пресноводных моллюсков с двустворчатой раковиной, одевающей тело с боков. Характерная особенность их — редукция головы. У большинства представителей имеется пара ктенидиев, превращенных в большие пластинчатые жабры. Некоторые Lamellibranchia могут достигать значительных размеров, а именно встречающаяся в Индийском и Тихом океанах Tridacna gigas имеет раковину до 1,35 м, масса ее может быть больше 250 кг. Строение и физиология. Внешняя морфология. Тело чаще всего продолговатое, более или менее сплюснутое с боков и билатерально сим- 472 метричное. Голова редуцирована, так что тело состоит из туловища и ноги. На переднем конце туловища лежит рот, на заднем — порошица. Между ними на брюшной стороне туловища выдается нога (рис. 463, 464). 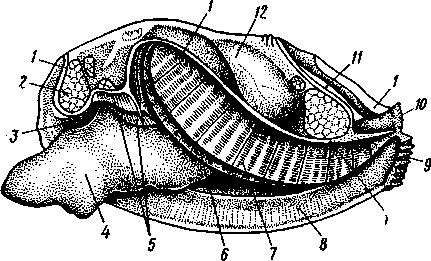 У немногих наиболее примитивных представителей (отряд Protobran-chia) нога, как и у Gastropoda, снабжена плоской ползательной подошвой (см. рис. 478). У всех прочих Lamellibranchia нога сильно сплюснута с боков и заострена по свободному краю, образуя подобие киля. Такая нога служит не столько для ползания, сколько для рытья песка или ила, в котором часто живут пластинчатожаберные. У некоторых форм, ведущих неподвижный образ жизни, нога становится рудиментарной (мидия — М у- tilus; см. рис. 475) или исче- Рис. 463. Анатомия беззубки Anodonta, раковина и левая мантия удалены (по Гауэс): СОВСеМ (устрица USt- / — линия, по которой обрезана мантия, 2 — передний Гей', СМ. РИС. 474) У МНОГИХ мускул-замыкатель, 3 — рот, 4 — нога, 5 — ротовые ло- г - ' пасти, 6 — левая внутренняя пояужабра, 7 — левая на- ПЛаСТИНЧаТОЖаберНЫХ На ружная полужабра, « — правая мантия, 9 — вводной си- НИЖН6Й ПОВерХНОСТИ НОГИ В фон- W-выводной сифон, а//д-задняя кишка, 12 - пе- особом вдавлении открывается так называемая биссу- совая железа. Она выделяет тягучие нити секрета биссуса, которые в воде быстро твердеют. При помощи получающихся прочных шелковистых нитей животные могут прикрепляться к подводным предметам (Му-lilus, Dreissena). Тело покрыто мантией (см. рис. 463); последняя свешивается с боков в виде двух больших мантийных складок. Между складками и телом остается полость — мантийная, в которой помещаются нога и жабры. На спине мантийные складки переходят одна в другую, а на передней, брюшной и задней сторонах обычно заканчиваются свободным краем, на котором иногда могут развиваться маленькие щупальца и даже глаза. 473 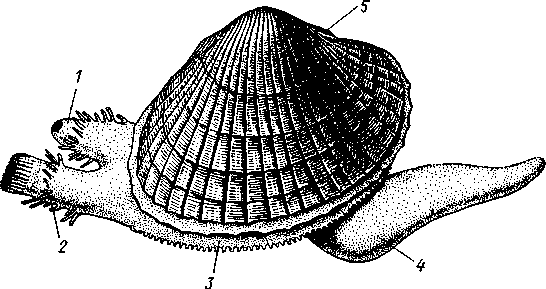 Рис. 464. Строение сифонов у зарывающегося в грунт моллюска Cardium edule (по Мейеру и Мебиусу): 1 — выводной (клоакальный) сифон. 2 — вводной (жаберный) сифон, 3 — мантия, 4 — нога, 5 — раковина 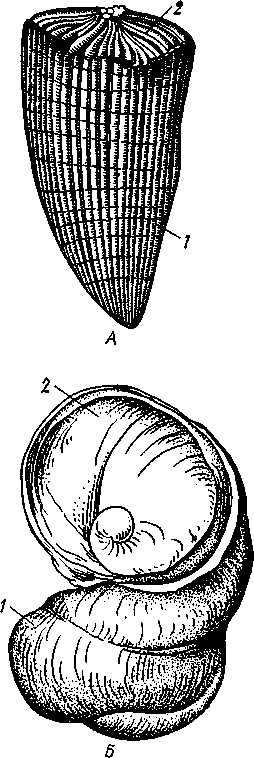 Нередко, однако, края обеих складок могут на известном протяжении срастаться, образуя при этом от 2 до 4 отверстий, через которые мантийная полость сообщается с внешней средой. Чаще всего срастание происходит по заднему краю мантии в двух участках, в результате чего общая мантийная щель распадается на три отверстия: два небольших задних и одно большое, ограниченное передними и брюшными участками мантийных складок. Нижнее из двух задних отверстий служит для введения в мантийную полость воды, содержащей пищевые частицы и служащей для дыхания, и называется вводным сифоном, верхнее — для выведения из нее воды и экскрементов — это выводной сифон. Большое передне-брюшное отверстие позволяет ноге высовываться из мантийной полости наружу. У форм, глубоко зарывающихся в ил или песок, края вводного и выводного сифонов часто вытягиваются в длинные мускулистые трубки (рис. 464). Выдаваясь над поверхностью грунта, они обеспечивают доступ свежей воды в мантийную полость. Наружным эпителием мантийных складок выделяются обе створки раковины-. Створки прикрывают тело с боков, и у большинства Lamellibranchia развиты одинаково. Однако у некоторых форм они различны главным образом вследствие прирас-тания животного одной стороной к морскому дну. Так, у устриц (см. рис. 474) прикреплена ко дну левая створка; она крупнее 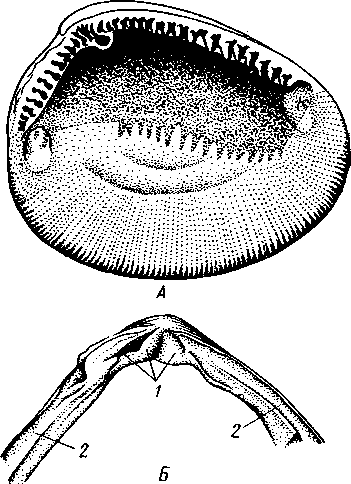 Рис. 465. Раковины рудистов Hippurites (А)(из Циттеля) и Requlenla (Б): 1 — створка, вмещающая тело животного, 2 — створка, играющая роль крышечки Рис. 466. Типы замков пластинчатожаберных. А — равнозу-бый замок; Б — разнозубый замок (из Ремане): 1 — главные зубы, 2 — второстепенные зубы 474 другой, более выпукла и вмещает все тело, тогда как правая; створка тонка и функционирует лишь в качестве крышечки. Особенно сильна разница между обеими створками у некоторых ископаемых форм (рис. 465); у рудистов, например, одна из створок имеет вид высокого конуса, а другая, по-видимому, служила крышечкой (Hip-purites). Некоторые пластинчатожаберные, например корабельный червь (Teredo; см. рис. 476), обладают сильно редуцированной раковиной, маленькие створки которой прикрывают лишь 1/20 часть тела. В типичных случаях обе створки выпуклы, причем наиболее выпуклые точки их лежат у спинного края раковины и называются макушками створок. Макушки являются наиболее старыми, начальными частями створок, к которым впоследствии присоединяются по краю все новые слои извести. Соответственно на раковинах можно различать ежегодные слои прироста, идущие параллельно свободному краю раковины и позволяющие определять возраст животного. На спинной стороне тела створки связаны между собой, во-первых, лигаментом, во-вторых, замком. Лигамент состоит из эластического вещества и соединяет обе створки в виде короткой поперечной ленты. Наружный слой лигамента непосредственно переходит в таковой створок, так что раковина, в строгом смысле слова, состоит из одного куска, перетянутого и перегнутого на спине. Вследствие своей эластичности лигамент держит обе створки полуоткрытыми. Замком называется соединение створок при помощи зубовидных отростков (зубов) спинного края, входящих в углубления противоположной створки. Различают 2 главных типа замков: равнозубый со значительным числом зубьев одинаковой величины и формы (Nucnla; рис. 466,Л; Area) и разнозубый с небольшим числом зубов разной формы (рис. 466,Б);первый тип зубов более древний. У некоторых форм (например, беззубки — Anodonta) замок может отсутствовать и тогда створки соединены лишь лигаментом. Для захлопывания раковины служат замыкательные мышцы, которых бывает две или одна. Они имеют вид толстых мускульных пучков, идущих поперек тела моллюска от одной створки к другой. Как в местах 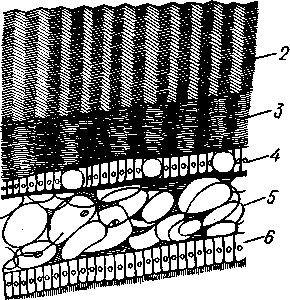 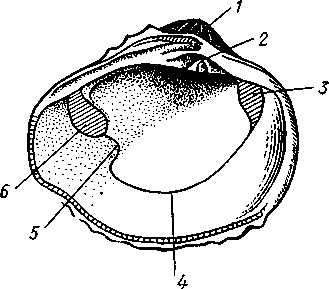 Рис. 467. Створка раковины с внутренней стороны (по Лангу): / — макушка раковины, 2 — спинной край с замком, 3 — отпечаток переднего мускула-замыкателя, 4 — отпечатки мышц, прикрепляющих край мантии к раковине (мантийная линия), 5 — «сифонный заливэ — место отхож-дения трубок сифонов, 6 — отпечаток заднего мускула-замыкателя Рис. 468. Разрез через раковину и мантию беззубки Anodonta (по Лейдигу): 1 — конхиолиновый слой, 2 —- фарфоро-видный, или призматический, слой, 3 — перламутровый слой, 4 — эпителий наружной поверхности мантии, 5 — соединительная ткань мантии, б — эпителий внутренней поверхности мантии 475 прикрепления мышц к створкам, так и по краю мантийных складок на внутренней поверхности раковины получаются легкие отпечатки, по присутствию или отсутствию которых можно судить о числе и расположении замыкательных мускулов, о степени развития сифонов и т. п. (рис. 467). Отпечатки хорошо сохраняются на ископаемых раковинах, что позволяет наряду с некоторыми другими данными судить о внутреннем строении давно вымерших животных. Наружный тонкий слой раковины (рис. 468), или периостракум, состоит из органического вещества, конхиолина, и нередко стирается на выпуклых частях створки (у макушки). Под ним залегает призматический, или фарфоровидный, слой, слагающийся из тесно прилегающих друг к другу призмочек углекислой извести, поставленных перпендикулярно поверхности раковины. Этот слой обладает значительной толщиной. Самый внутренний слой, перламутровый, образуется тончайшими, лежащими в несколько слоев известковыми листочками, между которыми залегают столь же тонкие прослойки конхиолина. В перламутре имеет место интерференция световых лучей; вследствие этого перламутр блестит и переливает разными цветами. Перламутровый слой подстилается эпителием мантии, который и выделяет раковину. Этот же эпителий у некоторых Lamellibranchia формирует жемчуг. Если какие-нибудь очень мелкие частицы, например омертвевшие клетки или зернистые продукты выделения, или тельца постороннего происхождения, например песчинки, нередко паразиты, попадут в промежуток между раковиной и эпителием маитии, то они все больше и больше обволакиваются концентрическими слоями перламутра и превращаются в жемчужины. Пищеварительная система (рис. 469). Рот расположен на переднем конце тела над основанием ноги. По бокам рта имеются 2 пары длинных треугольных ротовых лопастей. Они покрыты ресничками, подго- 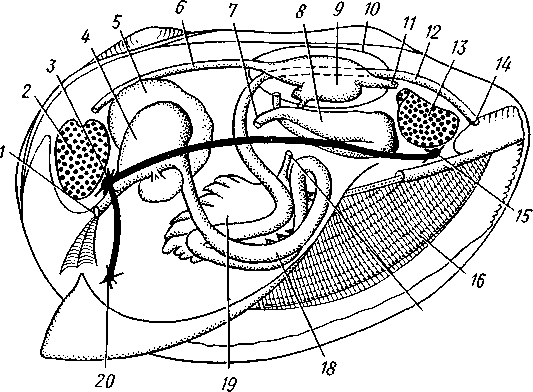 17 476 Рис. 469. Схема внутреннего строения пластинчатожаберного моллюска (из Реманс): / — рот, 2 — передний мускул-замыкатель, 3 — церебро-плевральный ганглий, 4 —желудок, 5 — печень, 6 — передняя аорта, 7 — наружное отверстие почки, « — почка, открывающаяся в перикардий, 9 — сердце, 10—перикардий, 11 — задняя аорта, 12 — задняя кишка, 13 — задний мускул-замыкатель. 14—анальное отверстие. 15 — висцеро-париеталь-ный ганглий, 16—жабры. 17 — отверстие гонады, 18 — средняя кишка, 19 — гонада, 20 — педальный ганглий няющими пищевые частицы к ротовому отверстию. Редукция головы вызывает атрофию тех частей кишечника, которые у прочих моллюсков помещаются в голове, а именно глотки, терки, челюстей и слюнных желез. Рот ведет прямо в короткий пищевод, который открывается в меш-ковидный желудок. Недалеко от места впадения пищевода, но более вен-трально, от желудка отходит средняя кишка. В заднюю часть желудка открывается отверстие слепого мешковидного выроста, в полости которого формируется прозрачный студенистый стержень — кристаллический стебелек. Он состоит из мукопротеинов и ферментов (амилазы, гликоге-назы и др.). Свободным концом стебелек вдается в просвет желудка, где постепенно растворяется, высвобождая пищеварительные ферменты, которые и обеспечивают первичную обработку пищи. По бокам желудка помещается парная, хорошо развитая печень, состоящая из множества мелких долек и впадающая своими протоками в желудок. Средняя кишка спускается от желудка в основание ноги, делает несколько изгибов и затем направляется по спинной стороне туловища к его заднему концу. Она переходит в заднюю кишку, которая обычно пронизывает желудочек сердца и заканчивается порошицей над задней замыкательной мышцей. Будучи малоактивными, подчас и неподвижно прикрепленными (например, устрица — Ostrea, мидия — Mytilus и др.) животными, пластинчатожаберные питаются пассивно. Пищей им служат мелкие взвешенные в воде частицы — детрит, планктонные организмы и бактерии, которые моллюски отфильтровывают из воды, проходящей через мантийную полость. 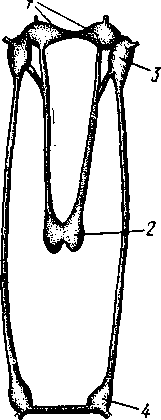 Нервная система (рис. 470) состоит из 3 пар ганглиев. Цереброплевральные ганглии представляют продукт слияния двух пар узлов; это доказывается тем, что у примитивных Protobranchia плевральные узлы еще несколько обособлены от церебральных. Цереброплевральные узлы соединяются над глоткой тонкой церебральной комиссурой. В ноге залегает пара педальных ганглиев, которые соединяются с церебро-плевральными посредством двух длинных коннекти-вов. Еще более длинные коннективы идут от церебро-плевральных узлов к паре висцеропариетальных ганглиев, лежащих под задним мускулом-замыкателем. Эти ганглии кроме внутренностей иннервируют ос-фрадии и жабры. Органы чувств развиты слабо, чему, по-видимому, служит причиной малоподвижный роющий образ жизни. У основания жабр имеются осфрадии, а в соседстве с педальными ганглиями всегда находятся два ста-тоциста. Рис. 470. Схема нервной систем и Lamellibranchia (по Гессе): 1 — церебральные ганглии, 2 — педальные ганглии, 3 — плевральный ганглий, 4 — висцерона]^е-тальный ганглий Головные щупальца и глаза, гомологичные соответствующим образованиям брюхоногих, отсутствуют. Имеются случаи, когда типичные ограны зрения вторично возникают или по всему краю мантии (гребешок— Pecten), пли по оторочке сифонов (сердцевидка— Cardium). У гребешка мантийный край несет свыше сотни отдельных инвертированных глаз довольно сложного строения. Органами осязания пластинчатожаберных служат отчасти околоротовые лопасти, а также различные 477 щупальцевидные придатки, развивающиеся по свободному краю мантии (Pecten) или по краям сифонов. Органы дыхания (см. рис. 463) представлены рядом модификаций типичных ктенидиев. У представителей примитивного отряда Protobran-chia по бокам основания ноги в мантийной полости лежит по двояко-перистому ктенидию, состоящему из общей оси и двух рядов треуголь- 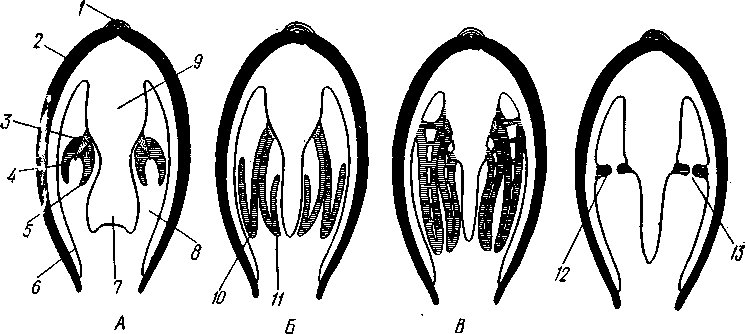 Г Рис. 471. Жабры Lamellibranchia. Схематические поперечные разрезы через тело в области жабр. А — Protobranchia; Б — Filibranchia; В — Eulamcllibranchia; Г—Septi- branchia (из Ланга): / — лигамент, 2 — створка раковины, 3 — ось ктенидия, 4 — наружный лепесток ктенидия, 5 — внутренний лепесток ктенидия, 6—мантия, 7 — нога, 8 — мантийная полость, 9 — туловище, 10—наружная нить, состоящая из нисходящего и восходящего колен, 11 — внутренняя нить, 12 — мускулистая перегородка, произошедшая за счет участков жабры, прирастающих к мантии и ноге, 13 — отверстие в перегородке лых боковых жаберных лепестков (рис. 471). Одним краем оси каждый ктенидий прирастает к потолку мантийной полости, а концы лепестков несколько выдаются в полость мантии. У нитежаберных (Filibranchia) лепестки обоих рядов необычайно удлиняются и принимают вид жаберных нитей, настолько длинных, что каждая нить, свешиваясь книзу, образует сначала нисходящее колено, а затем загибается кверху и дает восходящее колено. У внутреннего ряда лепестков восходящее колено направлено к ноге, у наружного — к мантийной складке (рис. 471,Б).У части Filibranchia жаберные нити свободны. У других представителей они связаны соединительноткаными мостиками. У отряда Eulamellibranchia, наконец, соединение между жаберными нитями становится еще более полным. В результате каждая половина жабры превращается в двухслойную решетчатую пластинку (рис. 471.В). Таким образом, каждая из четырех жабр Lamellibranchia на самом деле соответствует лишь одной половине настоящего ктенидия. Эпителий жабр во многих местах имеет мерцательный характер. У небольшого отряда Septibranchia ктенидий атрофируются, а в мантийной полости образуется мускульная горизонтальная перегородка, делящая полость на два участка: нижний и верхний, или дыхательный (рис. 471,Г); в последнем и происходит газообмен. Кровеносная система. Сердце пластинчатожаберных помещается на спинной стороне тела и лежит в тонкостенной околосердечной сумке (перикардий). Как показывает история развития, зачаток сердца закладывается парным и у некоторых низших Lamellibranchia, а именно у Area животное обладает двумя сердцами. У прочих пластинчатожабер- 478 ных оба зачатка сливаются и образуют непарное сердце, состоящее из желудочка и двух предсердий. У примитивных форм отряда Protobran-chia слияние происходит над кишкой. У большинства Lemellibranchia правый и левый зачатки охватывают заднюю кишку и сливаются под и над нею, следствием чего и является пронизывание желудочка задней кишкой (см. рис. 469) — черта, характерная для пластинчатожаберных. От желудочка берут начало два мощных артериальных сосуда — передняя и задняя аорты. Передняя аорта идет над кишкой вперед и отдает от себя артерии к внутренностям, ноге и передней части мантии. Задняя аорта следует назад под кишечником и вскоре распадается на две задние мантийные артерии. Из артерий кровь попадает в систему лакун в соединительной ткани и, наконец, собирается в большую продольную венозную лакуну, лежащую под перикардием. Из лакуны кровь направляется в проходящий вдоль основания каждой жабры приносящий жаберный сосуд, проникает отсюда в жаберные нити, окисляется и возвращается в выносящий жаберный сосуд, проходящий параллельно приносящему. Выносящие жаберные сосуды сообщаются с предсердиями сердца, из которых кровь проходит в желудочек (см. рис. 469). Выделительная система состоит из пары почек, которые лежат в задней половине тела по бокам и несколько ниже кишки. Они имеют вид двух обширных трубчатых мешков с железистыми стенками. Каждый мешок сложен по длине-вдвое так, что принимает V-образную форму с углом, обращенным назад. Обе передние ветви заканчиваются отверстиями; одним из них почка сообщается с перикардием, а другим — с мантийной полостью. По своему происхождению почки пластинчатожаберных — типичные целомодукты. / Известное участие в выделении принимают и стенки перикардия. Клетки передней половины перикардия имеют железистый характер и образуют перикардиальные железы. Последние иногда обособляются от остального перикардия в виде двух мешков — кеберовых органов, сообщающихся с ним отверстиями. Продукты выделения этих желез попадают в перикардий, а оттуда выводятся через почки наружу.   |