В.А. Догель - Зоология беспозвоночных. Тип губки (spongia, или porifera)
 Скачать 15 Mb. Скачать 15 Mb.
|

|
/— Оуккальный ганглий, 1 — педальный ганглий, 3 — оптический нерв, 4 — висцеральный ганглий, 5 — церебральный ганглий, 6 — нерв, идущий к капюшону и щупальцам, 7 — нервы рук, 8 — место отхождения оптического нерва, 9 — мантийный нерв, 10 — нерв, идущий к внутренностям, 11 — нерв воронки, 12 — инфундибулярный ганглий, 13 — брахиальный ганглий, (12и 13 — производные педального ганглии) 492 общается с внешней средой (рис. 489, Л). У всех прочих Cephalopoda глаза носят характер очень крупных, замкнутых, сложно устроенных глазных пузырей (рис. 489, 6). Строение их легче всего понять, прослеживая историю развития глаза. Сначала у зародыша образуется первичная глазная ямка (стадия Nautilus), которая отшнуровывается от кожи и дает глазной пузырь. Глубокая часть стенки пузыря превращается в 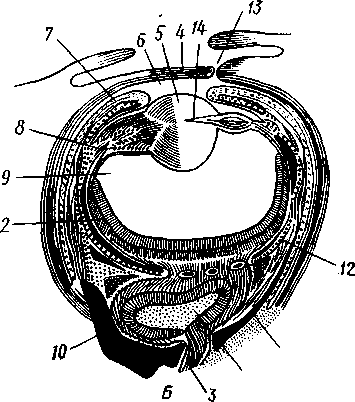 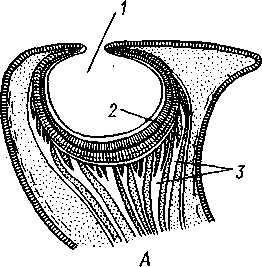 10 11 Рис. 489. Глаза головоногих. А — разрез глаза Nautilus (Tetrabranchia) (из Гешелера); Б — разрез глаза Sepia officinalis (Dibranchia) (по Генсену, с изменениями): 1 — полость глазной ямки, сообщающейся с внешней средой, 2 — сетчатка, 3 — зрительный нерв, 4 — роговица, 5 — хрусталик, 6 — передняя камера глаза, 7 — радужина, 8 — ресничный мускул, при сокращении приближающий хрусталик к сетчатке, 9 — стекловидное тело, 10 — глазные отростки хрящевой капсулы, // — оптический ганглий, 12 — склера (хрящевая оболочка глаза), 13 — наружное отверстие камеры глаза, /4 — эпителиальное тело сетчатку, тогда как наружная, прилегающая к коже часть дает в совокупности с наружным эпителием двухслойное эпителиальное тело. Вокруг глаза появляется кольцевая складка кожи — радужина. Она нарастает над глазом в виде свода, но не вполне смыкается, так что в центре свода остается отверстие — зрачок. Между тем обоими слоями эпителиального тела выделяется одним слоем кнаружи, другим — в полость глазного пузыря по прозрачному полушарию. Совокупность обоих полушарий дает шаровидный хрусталик, перерезанный тонкой двойной пластинкой эпителиального тела. Тем временем на глаз нарастает новая кольцевидная складка кожи — роговица. У большинства головоногих свод, образуемый роговицей над радужинным сводом, не вполне смыкается, сохраняя эксцентрическое отверстие. Сетчатка глаз состоит из очень длинных (иногда до 0,5 мм) клеток; к ней подходит мощный зрительный нерв, образующий под самым глазом очень крупный зрительный ганглий из нескольких ярусов нервных клеток. Глаза головоногих обладают аккомодацией, которая совершается, однако, не изменением кривизны хрусталика (как у человека), а его приближением или удалением от сетчатки; для этого служит особый ресничный мускул, прикрепленный к экватору хрусталика. Кроме того, в глазу Cephalopoda имеются приспособления к видению при более сильном и более слабом освещении. 493 Острота зрения животного отчасти зависит от числа воспринимающих свет элементов. В глазу головоногих этих элементов очень много. Так, у каракатицы Sepia на 1 мм2 сетчатки приходится 105000 клеток; у кальмара Loligo — даже 165000. Все это убеждает нас в большой сложности строения глаз головоногих, которые в этом отношении не уступают даже глазам позвоночных. У многих головоногих (особенно у глубоководных) в коже имеются особые органы свечения (см. рис. 480,Г),по строению несколько похожие на глаза. Так, у Lycoteuthis diadema 22 подобных органа, из которых часть светится ультрамариновым, часть — светло-голубым, а некоторые— рубиново-красным фосфорическим светом. Органы дыхания. Головоногие имеют настоящие ктенидии. У всех Dibranchia их 2, у Tetrabranchia (Nautilus) —4. Ктенидии расположены в мантийной полости симметрично по бокам туловища (см. рис. 491, 482). Жабры двоякоперистые, свободными концами направлены вперед. Обмен воды в мантийной полости обеспечивается сокращением мантийных мышц и деятельностью воронки. По количеству жабр Cephalopoda делятся на два подкласса: четырехжаберные (Tetrabranchia) и дву-жаберные (Dibranchia). Кровеносная система. Сердце всех головоногих состоит из одного желудочка, в который открываются с боков два (подкл. Dibranchia; рис. 490) или четыре (Tetrabranchia) предсердия. От переднего и заднего концов желудочка отходят две аорты: головная и внутренностная. Головная аорта идет рядом с пищеводом и дает ветви к голове и щупальцам. Внутренностная снабжает кровью кишечник и половые органы. Артерии разбиваются на сеть капилляров, из которых берут начало вены.- Вены рук собираются в голове в одну кольцевую вену, от кото- 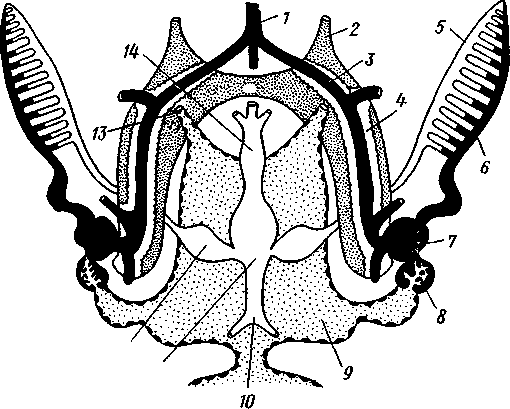 12 77 494 Рис. 490. Центральная часть кровеносной системы, жабры и почки каракатицы Sepia (из Кестнера): / — головная вена, 2 — наружное отверстие почек. 3—полая вена, 4—почка, 5 — уносящий жаберный сосуд, 6 — приносящий жаберный сосуд, 7 — венозное (жаберное) сердце, 8—перикардиальная железа, 9 — перикардиальная полость, 10 — внутренностная аорта, ;/ —желудочек, 12—предсердие, 13 — реио-перикардиальное отверстие, 14 — головная аорта (черным обозначены сосуды, по которым циркулирует венозная кровь, белым — сердце и сосуды с артериальной кровью)  5--М  рой рядом с головной аортой, но назад, идет крупная головная вена. Головная вена (рис. 490) делится на заднем конце на 2 или 4 (смотря по числу жабр) приносящих жаберных сосуда (полые вены), которые принимают еще кровь из мантии и несут венозную кровь к жабрам. Перед вступлением в жабры они образуют сократимые мускулистые расширения, так называемые венозные (жаберные) сердца, которые ритмическими сокращениями способствуют поступлению крови в жабры. Кровь окисляется в капиллярах жабр и затем по уносящим сосудам изливается в предсердия сердца. Капилляры вен и артерий в коже и мышцах непосредственно переходят друг в друга, лишь в некоторых местах между ними остаются лаку-нарные пространства, что делает кровеносную 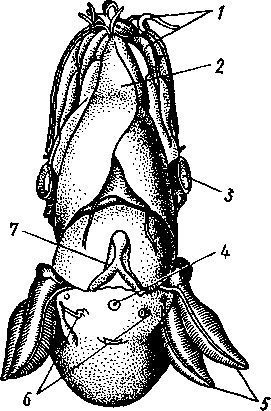 Рис. 491. Мантийный комплекс Nautilus. Вид животного, вынутого из раковины, с брюшной стороны, мантия удалена (по Тиле): / — щупальца, 2 — воронка, 3 — глаз, 4—> анальное отверстие, 5 — жабры, 6 — экскреторные отверстия, 7 — пенис Рис. 492. Сперматофор каракатицы Sepia (по Мильн-Эдвардсу): / — закрученный конец спер-матофора, 2 — различные части семяизвергательного аппарата, 3 — соединительный тяж, 4—хитиноидная оболочка, 5 — резервуар с семенем систему головоногих почти замкнутой. Кровь содержит гемоцианин — богатое медью соединение, физиологически соответствующее гемоглобину крови позвоночных; на воздухе синеет. Выделительная система состоит из 4 (подкл. Tetrabranchia; рис. 491) или 2 (подкл. Dibranchia; см. рис. 490) почек. Наружные отверстия их лежат по бокам порошицы, на особых сосочках (см. рис. 482), внутренние концы почек открываются, как всегда, в перикардиальный отдел це-лома. Почки представляют обширные мешки, иногда (отр. Decapoda) срастающиеся друг с другом по срединной линии тела. Проходящие в тесном соседстве с почками приносящие жаберные сосуды (с венозной кровью) образуют многочисленные бахромчатые 495 слепые выпячивания (венозные придатки), которыми вдаются в стенки почек, этим облегчается извлечение почками продуктов обмена веществ из крови. Половая система. Головоногие моллюски раздельнополы, иногда с резким половым диморфизмом. Так, у Argonauta самец гораздо меньше самки (см. рис. 486). Половая железа непарна и залегает в задней части туловища, в половом участке целома. Половые клетки скапливаются в целоме и выводятся через половые протоки. Протоки первично парны, хотя у многих головоногих остается только левый проток. Парность протоков сохраняется у самцов и у самок Nautilus, у Octopoda и части Decapoda. Протоки обладают сложным и варьирующим строением. У самца ка« ракатицы Sepia имеется семяпровод, расширяющийся в семенной пузырек, железистый эпителий которого служит для образования спермато-форов. За пузырьком семяпровод снова суживается и переходит затем в широкую сперматофорную сумку, которая и открывается сбоку от порошицы половым отверстием. Живчики головоногих склеиваются в окруженные плотной оболочкой пакеты, или сперматофоры (рис. 492). Спер-матофоры наполнены живчиками и имеют особый канал для их выхода, заткнутый сложно устроенной пробочкой. Женские протоки состоят из короткого яйцевода с впадающей в него небольшой яйцеводной железой. Кроме того, . независимо от полового  отверстия, но по соседству с ним в мантийную полость открываются выводные каналы двух парных и одной непарной нидаменталь-ных желез, секрет которых служит для образования яйцевых оболочек. Рис. 493. Самец осьминога Ocythoe catenulata с гектокоти- лизированной рукой (по Пель- знеру): I — концевая нить гектокотнля, 2 — мешочек гектокотиля, внутри которого развивается нить. 3 — гектоко-тилизированная рука, 4 — воронка, 5 — руки Оплодотворение чаще всего происходит в мантийной полости самки. Роль копуля-тивного органа играет одно из щупалец, у самцов оно более или менее сильно отличается от остальных иным развитием присосок и по присутствию на нем специального ложкообразного придатка (рис. 493). При помощи этого щупальца самец вводит сперматофоры в мантийную полость самки. Особенно замечательным устройством обладает половое щупальце у самца Argonauta (см. рис. 486) и некоторых других близких к нему родов. Щупальце сначала образуется внутри особого кожистого мешка, в котором оно свернуто в спираль. Конец полового щупальца вытянут в длинную нить. Щупальце содержит полость, сообщающуюся с внешней средой двумя отверстиями: одним у основания, другим на конце. Когда щупальце вполне сформируется, то облекающий его мешок лопается и оно расправляется. Полость щупальца (еще не выяснено, каким образом) наполняется сперматофорами. При спаривании половое щупальце Argonauta отрывается, надолго сохраняет способность к движению и проникает целиком в мантийную полость самки, где оно вводит сперматофоры в женское половое отверстие. 496 Когда оторвавшиеся щупальца самцов были впервые найдены в манти-й-ной полости самок, они были приняты за паразитов, и Кювье дал им родовое название Hectocotilus; это название и до сих пор сохранилось за половым щупальцем головоногих, которое называют «гектокотилизиро-ванным». Развитие. Эмбриональное развитие Cephalopoda отличается многими особенностями, определяемыми главным образом богатством яиц желтком. Он занимает почти все яйцо, его нет только на анимальном полюсе. Дробление вследствие обилия желтка частичное, причем делится на бластомеры лишь анимальный полюс; здесь образуется округлая пластинка, или диск, зародышевых клеток (частичное дробление головоногих называется дискоидальным). Развитие зародыша идет главным образом за счет анимального диска, на котором зародыш расположен так, что головным концом он обращен к желтку, а задним торчит свободно. Желток обрастается эктодермой и мезодермой зародыша, но остается довольно резко обособленным от последнего в виде желточного мешка. По мере роста зародыша желток потребляется, и желточный мешок уменьшается в размерах. Щупальца закладываются позади рта в виде двух боковых рядов, воронка имеет вид двухлопастного, открытого на брюшной стороне образования и лишь позднее срастается в трубку. Все развитие протекает внутри оболочки яйца. Палеонтология. Наиболее древними считаются известные с кембрия Nautiloidaea — родичи современного наутилуса. У первых наутилоидей раковина еще не была спирально закрученной и имела вид конической трубки, поделенной на камеры и пронизанной узким сифоном (Orthoce-ras). Такие или слегка согнутые наподобие рога раковины (Cyrtoceras) были особенно многочисленны в ордовике. Позднее появились вытеснившие их виды с раковиной, скрученной спирально, как у современного наутилуса, сохранившегося с триаса до наших дней. В ордовике были распространены гигантские Endoceras (представители особого отряда), прямая раковина которых, тоже поделенная на камеры, достигала 4—5 м длины. В широком сифоне, занимавшем около '/з диаметра раковины этих моллюсков, помещалась значительная часть органов животного. Эндоцерасы вели придонный образ жизни.  Рис. 494. Аммониты. А — Turrilites catenatus, редкая среди аммонитов форма закручивания раковины в коническую спираль (по Куку); Б — планоспиральная раковина Cardiacs-ras cordatum — хорошо сохранилась скульптура стенки раковины (по Бо« дылеаскому) 497 В девоне появляется небольшая группа относительно мелких головоногих с прямой или слегка согнутой раковиной — бактриты. По современным представлениям эти животные, просуществовавшие до конца палеозоя, дали начало двум большим ветвям развития головоногих — аммонитам, известным с девона, и белемнитам. Это чрезвычайно многочисленная и богатая пред-ствителями группа отрядов, существовавшая в палеозое и мезозое. В кайнозое аммониты отсутствуют, они вымерли в конце мелового периода. Аммониты обладали спирально закрученной многокамерной раковиной, обороты которой в большинстве случаев располагались в одной плоскости. Раковины аммонитов от- личались разнообразием формы (рис. 494), наружной скульптуры и размеров. Наряду с очень мелкими видами встречались настоящие гиганты, например Pachydiscus, планоспиральная раковина которого в диаметре достигала 2 м. Сведений о внутреннем строении аммонитов почти нет. По-видимому, они обладали вытянутым червеобразным телом. Полагают, что у них было 10 щупалец, чернильный мешок и, возможно, всего 2 ктенидия. Относятся ли аммониты к четырехжаберным или к двухжаберным, до сих пор еще не установлено. Вероятно, аммониты были хищниками и вели плавающий образ жизни. Они населяли моря земного шара на протяжении сотен миллионов лет. Бесчисленные остатки этих головоногих широко используются для определения геологического возраста слоев земной коры. Все рассмотренные группы головоногих обладали хорошо развитой раковиной, в которую целиком могло втягиваться тело животного. На этом основании палеонтологи объединяют их под названием наружнора-ковинных. Начиная с карбона, появляются головоногие, раковина которых снаружи обрастает мантией. Это внутреннераковинные, которые, как и аммониты, ведут начало от бактритов, но являются уже, несомненно, двужаберными. Эта ветвь головоногих достигла высшего расцвета в кайнозое. Сюда относятся современные кальмары и осьминоги. Среди ископаемых представителей этой группы особенно своеобразными были белемниты. Они появились в карбоне, достигли расцвета в юре и мелу и полностью исчезли к середине палеогена. Это были, по-видимому, плавающие моллюски. Задний конец их конической раковины (с. 489) имел вид массивного известкового конуса, иногда достигавшего значительной длины. Именно эта часть их раковины — рострум, наиболее часто встречается в мезозойских отложениях. Ископаемые головоногие по количеству видов несравненно многочисленнее, чем современные. Их насчитывают до 11 000 видов. Экология. Встречаясь исключительно в морях, головоногие ведут различный образ жизни. Большинство их относится к пелагическим животным. Такие виды, как Loligo, имеют торпедообразное, заостренное на заднем конце (который при движении направлен вперед) тело с хорошо развитыми плавниками. У некоторых донных осьминогов (Cirrothauma) между щупальцами развивается тонкая перепонка, так что животное принимает вид диска, лежащего на дне. Другие головоногие живут на дне, нередко прячась в расщелины между камнями. Глубоководные головоногие обладают иногда торчащими, наподобие телескопов, глазами. Головоногие — нередко весьма крупные животные. Самые мелкие из них измеряются несколькими сантиметрами, но на больших глубинах водятся настоящие гиганты. У подобных форм такая мускульная сила, что они очень редко попадаются в руки человеку, да и то лишь по счастливой случайности. Так, однажды удалось выловить извергнутые умирающим кашалотом во время агонии остатки съеденного им громадного головоногого Architeuthis; сохранившееся щупальце этого животного имеет длину около 10 м, следовательно, общая длина Architeuthis с вытянутыми ловчими щупальцами достигала 18 м. Практическое использование головоногих. Многие Cephalopoda служат предметом промысла. Каракатицы, кальмары и осьминоги употребляются в пищу в свежем, сушеном и консервированном виде. Их мясо по калорийности и вкусовым качествам не уступает говядине. Используется и секрет чернильного мешка каракатиц и кальмаров. После высуши- 498 вания и обработки едким калием осадок секрета дает акварельную краску сепию. Из этого же осадка приготовляют и натуральную китайскую тушь. Во всех странах мира ежегодно вылавливается около миллиона тонн головоногих моллюсков. Классификация. Современные головоногие делятся на два подкласса. ПОДКЛАСС 1. ЧЕТЫРЕХЖАБЕРНЫЕ (TETRABRANCHIA) Сюда принадлежат наиболее древние и примитивные формы, обладающие четырьмя жабрами, четырьмя предсердиями, четырьмя почками и значительным количеством щупалец; воронка в виде открытого желоба; раковина наружная, многокамерная. Че-тырехжаберные представлены несколькими группами отрядов (надотрядами), среди которых мы назовем лишь главнейшие. Надотряд 1. Наутилоидеи (Nautiloidea). Вымирающая группа, к которой в настоящее время принадлежит один род Nautilus — кораблик (см. рис. 481) с небольшим количеством видов, обитающих в Индийском и Тихом океанах. Ископаемые формы: Orthoceras. Надотряд 2. Аммониты (Ammonoidea). Группа, целиком вымершая в меловом периоде. Представители: Turrilltes, Ammonites (рис. 494). Внутреннее строение аммонитоп плохо известно, поэтому не ясно их систематическое положение. Устройство раковины приближает их к четырехжаберным. ПОДКЛАСС II. ДВУЖАБЕРНЫЕ (D1BRANCHIA) Головоногие, обладающие двумя жабрами, двумя предсердиями и двумя почками; воронка в виде трубки, раковина внутренняя, более или менее редуцированная. Отряд 1. Десятиногие (Decapoda). Имеют 10 щупалец, из них одна пара длинных, ловчих. Хорошие пловцы с вытянутым торпедообразным телом с боковыми плавниками. Представители: обыкновенная каракатица Sepia officinalis, гигантский кальмар Archi-teuthis, кальмар Loligo и др. (см. рис. 480). Ископаемые формы: Belosepia, Spirulirostra, К десятиногим относится вымершая в эоцене группа белемнитов (Belemnoidea) с прямой конической раковиной и массивным рострумом. Представитель: Belemnites. Некоторые авторы выделяют белемнитов в самостоятельный отряд Belemnitida. Отряд 2. Восьминогие (Octopoda). Большей частью донные формы с мешковидным телом и вполне редуцированной раковиной; щупалец 8, ловчих щупалец нет. Представители: аргонавт Argonauta, осьминоги Octopus и др. (см. рис. 480). Филогения типа Mollusca Происхождение типа моллюсков и их филогенетические связи с другими группами беспозвоночных животных были и остаются предметом оживленной дискуссии зоологов. Долгое время доминировала точка зрения, согласно которой моллюски в своем происхождении связаны с ан-нелидами. И действительно, в эмбриональном развитии моллюсков ряд признаков указывает на родство их с кольчатыми червями: спиральное детерминированное дробление, способ закладки мезодермы, личинка трохофора. В строении взрослых моллюсков сходны с кольчецами отношения между гонадами, целомом и целомодуктами. Более того, самые примитивные моллюски Amphineura и Monopla-cophora обнаруживают ясно выраженную метамерию, остатки которой в виде двух пар ктенидиев, почек и предсердий сохраняются у низших головоногих (Nautilus). Однако не все проявления метамерности могут приниматься во внимание. Расчленение раковины у Loricata и упорядоченное расположение мускульных пучков, так же как и большое количество жабр, вряд ли должны рассматриваться как пример сохранения моллюсками исходной метамерности. По мнению многих зоологов, это вторичное явление. Так, из большого и варьирующего у разных видов числа жабр только одна пара, расположенная около почечного отверстия и снабженная осфрадиями, вероятно, соответствует настоящим кте- 499 кидиям; остальные жабры возникли вторично для интенсификации функции дыхания. С аналогичным явлением мы сталкиваемся и у современных Monopla-cophora. Внешняя простота колпачковидной раковины и метамерное расположение мышц явно вторичны, так как наиболее древние ископаемые представители группы обладали спиральной раковиной и всего одной парой раковинных мышечных пучков. Значительно больший вес имеют следы метамерии в строении внутренних органов и в первую очередь органов, связанных с целомом. Наличие у Neopilina 6—7 пар целомодуктов, выполняющих функции почек и частично служащих для выведения половых продуктов, двух пар гонад и двух пар предсердий позволяет предполагать, что тело моллюсков первоначально состояло из небольшого числа (6—7) сегментов, унаследованных от далеких предков. Последними, по-видимому, были какие-то древние олигомерные кольчецы, тело которых состояло только из лар-вальных сегментов. Таким образом, тело современных моллюсков соответствует ларвальному телу кольчецов и вторично утратило метамерное строение. Уменьшение числа ряда образований (целомодуктов, предсердий, настоящих ктенидиев и т. д.) должно рассматриваться как результат олигомеризации гомологичных органов. Среди современных моллюсков наиболее примитивными, по-видимому, являются моноплакофоры. К особенно древним чертам организации Neopilina, несомненно, относятся: метамерия, двураздельность сердца и наличие продольных нервных стволов. По наличию двух желудочков сердца Monoplacophora сравнимы с зародышами пластинчатожаберных, брюхоногих и головоногих, у которых сердце сначала представлено парой зачатков, лежащих по бокам от кишки. При дальнейшем развитии зачатки сливаются, образуя один желудочек. У некоторых низших представителей класса Lamellibrar.chia (Area) слияния зачатков также не происходит, и взрослое животное имеет два сердца (с. 478). Таким образом, двураздельное сердце Monoplacophora представляет очень примитивную особенность, у большинства Conchifera проявляющуюся лишь в онтогенезе. У Amphineura непарное сердце. Из сказанного ясно, что кл. Monoplacophora во многих отношениях примитивнее современных панцирных. Естественно поэтому сделать вывод, что оба ствола моллюсков — Amphineura и Conchifera разделились очень рано. Monoplacophora еще очень близки к общему предку всех моллюсков. Однако последний, вероятно, отличался отсутствием настоящей раковины, имел еще невысокое туловище, сходное с таковым у Lori-cata, и, по-видимому, вел более активный образ жизни, нежели Neopilina. От этого гипотетического первичного моллюска берут начало две главные линии развития. Одна ведет к Amphineura, другая — к Conchifera. Среди боконервных наиболее примитивные черты: 8 спинных пластинок раковины, хорошо развитая радула, множественные жабры характерны для Loricata, тогда как Solenogastres, вероятно, представляют их вторично измененных потомков (утрата ноги и мантии, редукция раковины, радулы и т. д.). У самого корня ветви Conchifera находится класс Monoplacophora, объединяющий некоторые существенные признаки Lamellibranchia, Gastropoda и Cephalopoda. Кристаллический стебелек кроме Neopilina имеется у многих пластинчатожаберных и низших брюхоногих. Общее 500 сходство развития сердца у Neopilina и высших Conchifera уже отмечалось. На филогенетическую связь с Cephalopoda (Tetrabranchia) указывают остатки метамерии у Nautilus. Нервная система в виде продольных тяжей свойственна низшим Gastropoda и Nautilus. Таким образом, многое свидетельствует о происхождении высших классов Conchifera от форм, близких к Monoplacophorn. Однако современная Neopilina испытала уже некоторую специализацию, приспособившись к почти сидячему малоподвижному образу жизни. От примитивных Monoplacophora, вероятно, берет свое начало гипотетическая группа билатерально симметричных моллюсков (так называемые Prorhipidoglossa), соединявшая в себе, по мнению многих зоологов, наиболее примитивные черты строения современных Gastropoda и Lamellibranchia. От этой группы расходятся в виде веера классы Gastropoda, Lamellibranchia и Scaphopoda, постепенно удаляющиеся друг от друга в лице своих высших и наиболее специализированных представителей. Обособление брюхоногих базируется на развитии у них асимметрии, а эволюция пластинчатожаберных характеризуется главным образом редукцией головы и образованием двустворчатой раковины. Класс Cephalopoda стоит особняком, почему его приходится производить прямо от первичных моллюсков, близких к Monoplacophora, независимо от прочих классов Conchifera. Среди Cephalopoda подкласс че-тырехжаберных обладает, безусловно, более древними и примитивными чертами по сравнению с подклассом двужаберных. Существует, однако, и другая точка зрения на происхождение и филогению моллюсков. Ее сторонники считают, что моллюски берут начало непосредственно от плоских червей (кл. Turbellaria), а самыми примитивными формами являются бороздчатобрюхие (кл. Solenogastres), объединяемые в отр. Chaetodermatina (или Caudofoveata). Простота строения в этом случае оказывается первичной, а не возникает в результате вторичного упрощения. Соответственно моллюски должны рассматриваться как исходно несегментированные животные. Их целом, представленный главным образом перикардием и отделяющимися от него в процессе индивидуального развития гонадами, не гомологичен вторичной полости тела кольчатых червей и возникает в процессе эволюции независимо от последней. Увеличение же числа отдельных органов и упорядоченность их расположения — вторичное явление, которое можно рассматривать как псевдометамеризацию. Окончательное решение вопроса о природе моллюсков и их положении в системе животного царства, несомненно, требуют новых углубленных исследований. ТИП ЩУПАЛЬЦЕВЫЕ (TENTACULATA) Щупальцевые составляют группу вторичнополостных, олигомерных (малосегментных) животных не вполне выясненного происхождения. Щупальцевые ведут сидячий образ жизни, который оказал глубокое влияние на их организацию. Рассмотрим характерные признаки типа. 1. Тело неясно расчленено и слагается из трех сегментов. Первый — предротовая лопасть, или эпистом. Второй сегмент несет ротовое отверстие. Он окружен ресничными щупальцами, которые служат для подгона пищевых частиц ко рту и для дыхания. Щупальца сидят на щупаль-ценосце, или лофофоре, в основе имеющем форму подковы, или расположены по кругу. Третий сегмент — собственно туловищный. В ряде случаев эпистомальный сегмент вторично редуцируется. 501 2. Полость тела представлена целомом, который в соответствии с сегментацией разделяется на три отдела: передний эпистомальный, средний— сравнительно узкий, посылающий каналы в лофофор и щупальца, и задний, самый обширный — туловищный целом. Обособленность отделов целома может быть не всегда хорошо выражена. 3. Анальное отверстие лежит недалеко от рта, кишечник образует петлю. 4. Имеется кровеносная система, которая может вторично редуци-ваться. 5. Органы выделения Днефридии) представлены одной или двумя парами ресничных воронрк, открывающихся в целом. Являются ли эти образования метанефридиями или целомодуктами, пока окончательно не установлено. Иногда органы выделения редуцированы. 6. Половые железы образуются в туловищном целоме. 7. Дробление яйца полное, не спирального типа, из яйца развивается планктонная личинка, более или менее напоминающая трохофору. К типу Tentaculata относятся 3 класса: мшанки (Bryozoa), плечено-гие (Brachiopoda), форониды (Phoronidea). Кроме того, в качестве дополнения к типу мы помещаем класс внутрипорошицевых (Entoprocta), систематическое положение которых неопределенно. КЛАСС I. МШАНКИ (BRYOZOA) Класс мшанок содержит водных, преимущественно морских, реже пресноводных животных, ведущих сидячий образ жизни и образующих в результате бесполого размножения колонии. Чаще всего колонии бывают древовидными, ветвистыми. В некоторых случаях (Flustra) ветви колонии сплющиваются в одной плоскости и принимают характер пластинок, а сама колония становится похожей на пучок листьев. Если же колонии стелются по субстрату, то образуют на дне или на стеблях водорослей тонкие корочки. Каждая колония объединяет большое количество особей. Таким образом, по внешнему виду колонии мшанок больше всего напоминают гидроидных полипов. Размеры мшанок невелики: колонии измеряются несколькими сантиметрами, отдельные особи большей частью длиной не более 1 мм. Современных видов мшанок около 4000, ископаемых еще больше. Для понимания природы мшанок следует рассмотреть строение отдельной особи. Строение и физиология. Отдельные особи колонии мшанок (рис. 496) отчетливо подразделяются на два отдела, которые, однако, не являются сегментами. Передний конец особи свободно торчит в окружающую воду и несет на себе рот с венчиком щупалец вокруг. Задняя половина животного одета утолщенной кутикулой, производимой наружным эпителием тела, и имеет вид глубокой чашечки или мешочка. Эту часть животного нередко обозначают особым термином — цистид. Передняя половина мшанки лишена плотной кутикулы, нежная и при раздражении втягивается внутрь цистида; ее называют полипидом (рис. 497). Втягивание совершается при помощи двух особых мускулов-ретракторов. Они находятся по бокам кишечника и передним концом прикрепляются изнутри к стенке тела полипида по сторонам рта, а задним концом — к основанию цистида. При втягивании передней половины тела в цистид отдельные щупальца не вворачиваются, а весь венчик их уходит в глубь образующегося впячивання. Щупальца имеют вид длинных нежных выростов, в которые заходят каналы целома, и покрыты мерцательным эпителием; 502 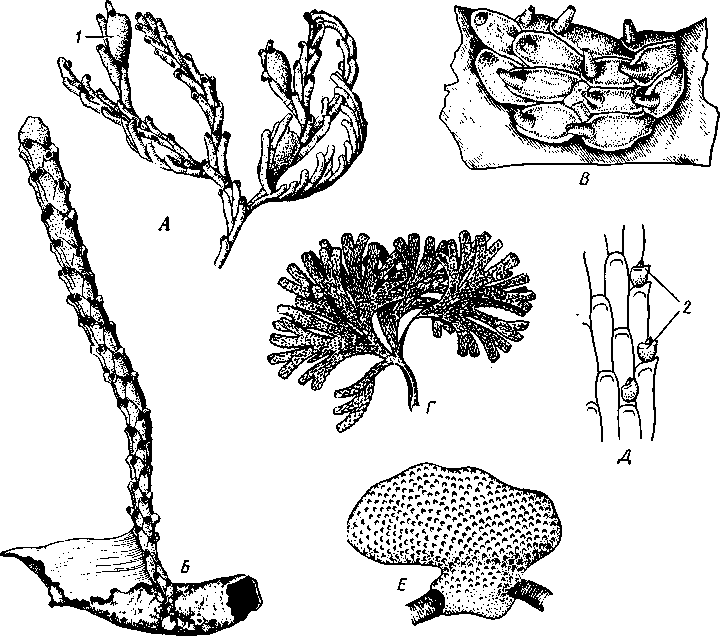 Рис. 495. Колонии морских мшанок подкласса Gymnolaemata. А — Idmonea tu-mida; Б — Crisia eburnea; В — Alcyonidium mamillatum; Г — Dendrobaenia flustrot-des; Д — участок колонии Dendrobaenia flustroides; E — Porella saccata: 1 — ГОНОЗОИД. 2 — авикулярия 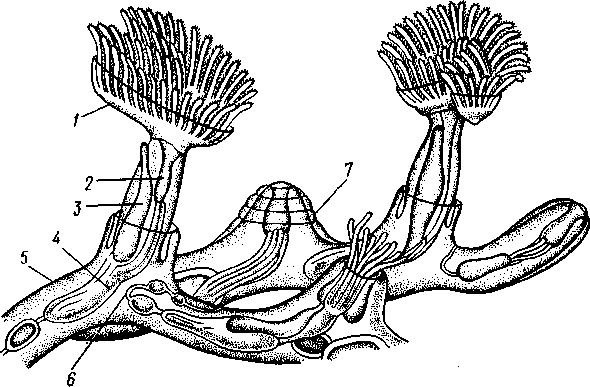 Рис. 496. Участок колонии Plumatella repens (из Матвеева)? / — полипид с расправленным лофофором, 1 — передний отдел кишечника, 3 — задняя кишка, 4 — желудок, 5 — стенка цистида, 6 — канатик со статобластамн. 7 — втянутый полипид 503 они служат, с одной стороны, для собирания пищевых частиц, с другой — в качестве органов дыхания. У пресноводных мшанок, образующих подкласс Phylactolaemata (Покрыторотые), щупальца в два ряда располагаются на особом щу-пальценосце — лофофоре, имеющем сверху, со стороны рта, форму под-ко.вы, у вершины которой и располагается ротовое отверстие (рис. 496, 497). Последнее прикрыто небольшим язычковидным выростом — эпи-стомом — предротовым отделом тела (рис. 497). Представители подкласса Gymnolaemata (см. рис. 495) (Голоротые мшанки) вторично утратили эпистом, а их щупальце вокруг ротового отверстия образуют простой венчик (рис. 498). Кутикула цистида может быть различной консистенции: в виде тонкой хитиновой пленки или остуденяется и превращается в толстый желеобразный слой, содержащий до 99% воды. В других случаях кутикула 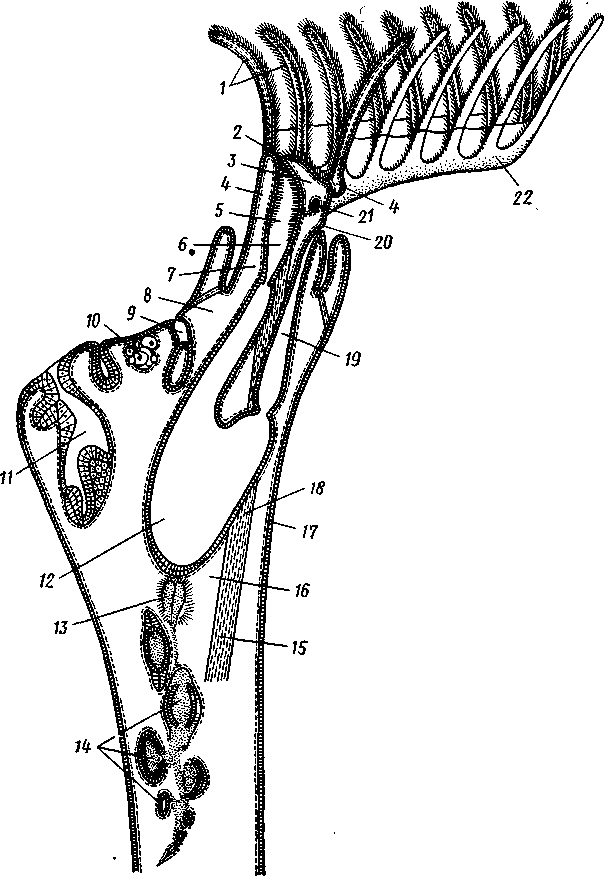 Рис. 497. Строение особи пресноводной мшанки из подкл. Phylactolaemata (из Ремане): I — щупальца, 2 — эпистом. 3 —* целом эпистома, 4 — целом ло-фофора, S—глотка, 6 — пищевод, 7 — полипид, 8 — цистид, 9 — оэций, 10 — яичник, 11 — почка, 12 — желудок, 13 — семенник, 14 — статобласт, 15 — канатик, 16 — туловищный целом, 17 — целомический эпителий, 18 — ретрактор, 19 — задняя кишка, 20 — анальное отверстие, 21 — ганглий, 22 — ло* фофор 504 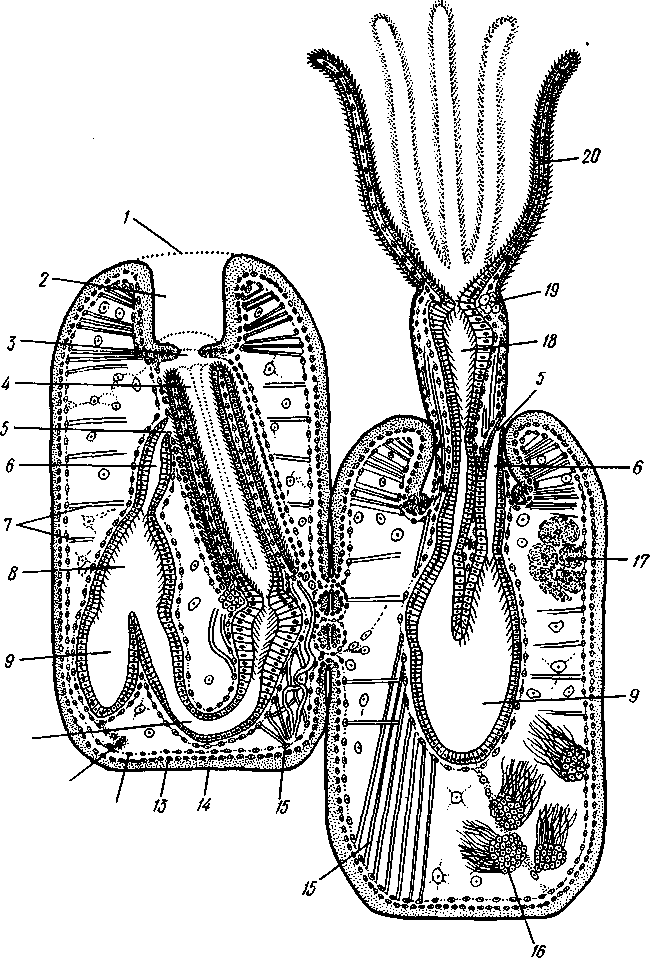 ID 11 12 505 I Рис. 498. Схематическое строение двух зооидов мшанок отряда Gymno-laemata (в расправленном и втянутом состоянии) (из Маркуса): / — отверстие цистида, 2 и 3 — части устьевой области цистида, 4 — венчик щупалец, втянутый внутрь цистида, 5 — анус, б — задняя кишка, 7 — мускульные волокна, пересекающие целом, 8, 9 — желудок, 10 — пищевод, // — канатик, 12 — перитонеаль-ный эпителий, 13 — стенка тела, 14 — кутикула, 15 — мускул-ретрактор, 16 — семенники, 17 — яичник, 18 — глотка, 19 — нервный узел, 20 — щупальца утолщается и пропитывается углекислой известью, становясь очень твердой. Наружный эпителий мшанок однослоен и у пресноводных форм (подкл. Phylactolaemata) подстилается двумя слабо развитыми слоями мышц: наружным из кольцевых и внутренним из продольных мышечных волокон. У морских голоротых мшанок (подкл. Gymnolaemata) мышечные слои отсутствуют. Слабое развитие кожно-мускульного мешка объясняется неподвижностью мшанок. Перитонеальный эпителий ограничивает обширную вторичную полость тела, в которой помещаются все внутренности. Полость тела (целом) тонкими перегородками делится на три отдела. Передний обладает небольшими размерами и залегает в эпистоме. Средний — кольцевой канал — окружает глотку и посылает слепые ответвления в щупальца. Задний, наиболее обширный, занимает почти все тело и называется туловищным целомом. Как уже говорилось, у голоротых мшанок эпистом вместе с целомом редуцируется. Пищеварительная система. Пищеварительный канал имеет характерную для многих сидячих животных подковообразную форму (см. рис. 496, 497, 498). Рот, лежащий внутри венчика щупалец (у подкл. Phylactotaemata— в средней части подковы лофофора, между двумя рядами ее щупалец), ведет сначала в небольшую глотку, а затем в длинную узкую трубку — пищевод. Пищевод переходит в мешковидный, V-образно изогнутый желудок; он впадает в одно из колен желудка, тогда как от другого колена отходит поднимающаяся кпереди тонкая кишка, открывающаяся наружу порошицей непосредственно позади венчика щупалец. Весь кишечник снаружи одет перитонеальным эпителием. От дна желудка отходит продолжение его перитонеальной выстилки в виде тонкой брыжейки, или канатика, тянущегося через полость тела назад и переходящего затем в наружный, пристенный слой перитонеума. Пища — мелкие одноклеточные водоросли, простейшие, коловратки и т. п. — подгоняется ко рту мерцанием ресниц на щупальцах. В глотке пища сбивается в пищевые комки и поступает через пищевод в желудок. Нервная система состоит из единственного надглоточного ганглия (см. рис. 497, 498), лежащего между глоткой и задней кишкой, от которого расходятся в разные стороны, но главным образом к щупальцам, периферические нервы. Единственными органами чувств являются особые осязательные волоски на наружной стороне щупалец. Интересно, что во время почкования ганглий почки возникает в виде ясного впячивания эктодермы, отшнуровывающегося от покровов; полость впячивания позднее исчезает, и ганглий становится массивным. Дыхание происходит через всю поверхность тела, но главным образом через щупальца. Кровеносной системы у мшанок нет. Отсутствие ее, вероятно, объясняется редукцией, вызванной малыми размерами тела, что, в свою очередь, связано с тем, что мшанки — животные колониальные. Выделительные органы у мшанок отсутствуют. Удаление продуктов обмена из организма осуществляется с помощью клеток-фагоцитов через стенки щупалец или стенку кишечника. Из последнего они выводятся вместе с экскрементами. У покрыторотых мшанок полости целомов соединяются трубчатыми образованиями, которые рассматриваются как видоизмененные целомо-дукты. Долгое время их считали органами выделения покрыторотых мшанок, но сейчас эта точка зрения оставлена. Половая система. Мшанки — гермафродиты. Половые клетки раз- 506 виваются под перитонеальным эпителием, на стенке тела или на канатике. Живчики образуют на канатике довольно большое скопление — семенник. Яйца формируются на внутренней стенке тела (рис. 498) в небольшом количестве, иногда по одному. Оплодотворение внутреннее — подвижные живчики выходят из одной колонии мшанок и активно проникают в другую, где в это время имеются созревшие яйца. Выход оплодотворенных яиц наружу совершается разными способами. У одних видов для этого используется специальная целомическая пора, расположенная между щупальцами и ротовым отверстием. Значительно чаще развивается специальный ресничный канал, который открывается наружу непосредственно между щупальцами. Наконец, как полагают, у ряда видов зрелые яйца обладают активным амебоидным движением и сами прокладывают себе дорогу через стенки родительской особи. У очень немногих видов оплодотворенные яйца выбрасываются в воду, где и осуществляется их дальнейшее развитие. Более обычно проявление у мшанок своеобразной «заботы о потомстве», когда яйца приклеиваются к покровам или даже поступают в особые образования — оэции (см. ниже), в которых протекает их развитие до полного формирования личинки. Колонии. У мшанок, как и у многих других колониальных организмов, наблюдается морфологическая и функциональная дифференци-ровка особей, образующих колонию. У сохранивших большое количество примитивных признаков покрыторотых мшанок в колониях представлены особи только двух типов. Основную массу составляют обычные индивиды. Кроме них имеются еще оэции, которые образуются за счет молодых почек, развитие которых рано останавливается. Почка у Phy-lactolaemata сначала представляет небольшое впячивание покровов, в полость которого и поступает яйцо. Оэции с развивающимся зародышем до конца сохраняет вид простого мешочка. В колониях подкл. Gymnolae-mata дифференциация особей выражена значительно сильнее. Оэции го-лоротых мшанок имеют вид выступающих бугорков, несущих полость, сообщающуюся с внешней средой. В этой полости протекает формирование личинки. У некоторых видов отдельные особи колонии резко увеличиваются в размерах и принимают характерную кувшинообразную форму. Они называются гонозоидами (см. рис. 495). В полости гонозоидов яйца претерпевают сложное развитие, включающее полиэмбрионию (с. 509). Ряд особей колонии специализируется для выполнения защитных функций. Это в первую очередь авикулярии (рис. 499). Последние имеют вид, сходный с птичьей головой (с шеей и мощным клювом). «Голова» представляет собой цистид, измененный и вытянутый в неподвижный отросток, отвечающий верхней части «клюва». Кроме того, на «голове» сидит подвижный кутикулярный крючок, отвечающий нижней части клюва; он способен то прижиматься к неподвижному отростку, то отводиться от него. Подвижный кутикулярный крючок есть результат видоизменения крышечки, имеющейся у многих мшанок на переднем крае цистида и захлопывающейся при втягивании полипида. Авикулярии захватывают мелких животных, заползающих на колонию, и убивают или прогоняют их. Разновидностью авикулярии являются вибракулы, которые отличаются отсутствием неподвижного выроста цистида, тогда как кутикулярный крючок вытягивается в длинный подвижный жгут. Размножение. Мшанки имеют половое размножение, но, кроме того, отличаются большим разнообразием бесполых способов размножения. Прежде всего образование колоний — результат не доведенного до конца бесполого размножения посредством почкования. Детали процесса 507 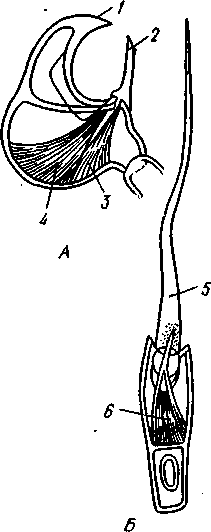 почкования у мшанок довольно сложны, но в общем он состоит в том, что на поверхности колонии образуются в определенных (у разных родов кл. Bryozoa различных) местах бугорки — зачатки будущих почек. Бугорок растет, и перетягивается у основания, но полностью не отделяется; почка остается в связи с колонией. На почке путем впячивания отдельных участков ее эктодермы закладываются кишечник и ганглий. Щупальца закладываются тоже в виде впячиваний, которые потом выворачиваются наружу. Соответственно месту образования почек на материнском организме, количеству их и дальнейшему способу почкования колония мшанок принимает различную форму. Почки формируются во вполне развитые особи и, в свою очередь, начинают размножаться почкованием. В результате такого наружного почкования происходит постепенный рост колонии.  Рис. 499. Авикулярия (А) и вибракула (Б) мшанки (из Догеля): / — неподвижная ветвь, 2 — подвижная ветвь, 3, 4—мышцы, раскрывающие и закрывающие «клюв», 5 — подвижный бич вибракулы, приводимый в действие мышцами (6) Рис. 500. Статобласт Crista-' tella mucedo. Вокруг цент-' ральной массы видно плава-• тельное кольцо, состоящее > из хитиновых камер, наполненных газом, и хитиноид-ные крючочки (по Догелю) Наряду с наружным почкованием у пресноводных мшанок (отр. Phylactolaemata) имеется еще особый способ внутреннего почкования при помощи образования статобластов (см. рис. 497; 500). Статоблас-ты — особые чечевицеобразные многоклеточные тельца, одетые плотной оболочкой; образуются преимущественно осенью. Зачаток статобласта появляется внутри канатика в виде кучки мезодермальных клеток. Одновременно с этим группа эктодермальных клеток мигрирует с поверхности тела внутрь канатика. Здесь за их счет формируется двухслойная эпителиальная оболочка, окружающая группу мезодермальных клеток. Эпителий выделяет на своей поверхности тонкую, но плотную скорлупу, состоящую из двух слоев, между которыми располагается слой воздухоносных камер. У некоторых видов по краю чечевицы образуется хитиновое кольцо с крючками, или зацепками. Статобласты освобождаются при разрушении материнского тела (например, зимой при отмирании колонии) и попадают наружу. Воздухоносные камеры позволяют им плавать в воде, а своими крючками они зацепляются за водоросли, неровности дна и т. п. Статобласты остаются зимой в покоящемся состоянии, 508 весной же оболочки их лопаются, клеточное содержимое прикрепляется ко дну и образует при помощи наружного почкования новую колонию. Легко видеть, что внутреннее почкование мшанок имеет большое сходство и одинаковое биологическое значение с образованием геммул у пресноводных губок (с. 108), Наконец, у некоторых мшанок, относящихся к подкл. Gymnolaemata, зимуют особые «зимние» наружные почки: они представляют собой одетые плотной оболочкой отдельные цистиды. От статобластов они легко отличаются по присутствию в них зачатка кишечника, мышц и половых клеток, тогда как внутренность статобласта занята сплошной массой мезодермальных клеток. Развитие. Все мшанки обладают половым размножением. Дробление яйца полное и во многих случаях почти равномерное. В результате возникает бластула, часто сплющенная в одном направлении. В этом случае она сначала имеет вид пластинки, а позднее становится похожей на чечевицу. |