В.А. Догель - Зоология беспозвоночных. Тип губки (spongia, или porifera)
 Скачать 15 Mb. Скачать 15 Mb.
|

|
КЛАСС V. ГОЛОВОНОГИЕ (CEPHALOPODA)  Рис. 480. Различные головоногие (по Хуну). А—Amphitretus pela-gicus (глубоководный плавающий осьминог); Б — Benthoctopus pro-fundorum (осьминог); В — Doratopsis sagitta (планктонный кальмар); Г — Lycoteuthis diadema (глубоководный пелагический кальмар со светящимися органами); Д — Cranchia scarba (планктонный кальмарчик); Е — Cirrothauma murrayi (донный, живущий на мягком грунте осьминог); Ж — Loligo edulis (пелагический кальмар) 485 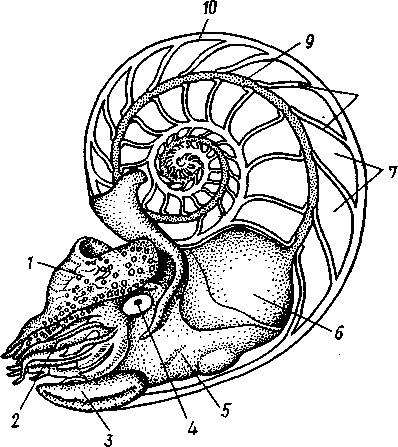 8 Рис. 481. Кораблик Nautilus pompilius (подкл. Tetrabranchia) с распиленной раковиной (по Оуэну): 1 — головной капюшон, 2 — щупальца, 3 — воронка. 4 — глаз, 5 — мантия, 6 — внутренностный мешок, 7 — камеры, 8 — перегородки между камерами, 9 — сифон, 10 — стенка раковины Класс головоногих (рис. 480) содержит около 700 живущих в морях видов наиболее сложно организованных, крупных, а иногда даже очень крупных (до 18м) моллюсков. Это свободноплавающие, реже ползающие, очень подвижные хищники, преимущественно обитающие в теплых морях. Тело их билатерально симметрично, с резким разделением на голову и туловище. Нога превращена в щупальца, или руки, которые вторично сместились на голову и окружают ротовое отверстие. Другая часть ноги представлена так называемой воронкой, лежащей на брюшной стороне у входа в мантийную полость. Раковина у примитивных форм наружная, многокамерная, у высших представителей — внутренняя, часто более или менее редуцированная. Нервная система и органы чувств достигают большого совершенства. Строение и физиология. Внешняя морфология. Голова несет на переднем конце рот, окруженный щупальцами. У немногих древних форм, например, кораблика Nautilus (рис. 481), имеется значительное и не вполне определенное количество тонких, червеобразных щупалец, которые служат\ для захватывания добычи и могут втягиваться в особые влагалища. Сильно разросшиеся влагалища дорзальных щупалец составляют так называемый капюшон — мускулистое образование, с помощью которого животное может закрывать вход в раковину. У высших головоногих всего 8 (отр. Octopoda) или 10 щупалец (отр. Dec.apoda), устроенных иначе, чем у наутилуса. Восемь щупалец Octopoda представлены широкими у основания, заостряющимися к концу мускулистыми выростами с продольным желобом на обращенной ко рту внутренней стороне. Эта сторона щупалец усажена многочисленными крупными дисковидны-ми присосками, позволяющими животному необычайно прочно присасываться ко дну и схватывать добычу. У Decapoda кроме этих 8 щупалец есть еще 2 гораздо более длинных ловчих щупальца, расширенных на конце. По бокам головы расположена пара очень крупных глаз. Туловище со всех сторон одето мантией. На спине она образует покровы самого туловища; на брюшной стороне она отделена от туловища мантийной полостью. На месте перехода туловища в голову мантийная полость сообщается щелевидным брюшным отверстием с внешней средой. Для замыкания брюшной щели у высших головоногих есть особое приспособление в виде пары полулунных ямок на брюшной стороне туловища, соответственно которым на внутренней поверхности мантии ле- 486 жат два твердых, укрепленных хрящом бугра, или запонки. Действием мантийных мышц мантия прижимается к туловищу, запонки входят в полулунные ямки и как бы пристегивают мантию к телу, закрывая этим брюшную щель. Между обеими запонковыми ямками на брюшной стороне туловища помещается воронка, мускулистая коническая трубка, приросшая к туловищу. Расширенный задний конец воронки открывается в мантийную полость, суженный передний смотрит наружу. Воронка служит для движения. Когда мантийная щель закрыта, мантия прижимается к туловищу действием многочисленных находящихся в ней мышц и вода из мантийной полости с силой выталкивается наружу через переднее отверстие воронки, сообщая животному обратный толчок. Затем щель раскрывается и полость мантии заполняется окружающей водой. Далее происходит новое замыкание щели, сжимание мантии и новый толчок. Ритмические сокращения мантии и выталкивание воды служат, с одной стороны, в качестве дыхательного акта для смены воды в мантийной полости, с другой — для плавательного движения, которое совершается у головоногих толчками задним концом вперед. По бокам тела нередко образуются кожные складки, поддерживающие тело в равновесии и служащие плавниками. В мантийной полости на брюшной стороне тела открывается порошица, по бокам ее расположены половые и почечные отверстия и ктенидии. Нога, на первый взгляд, отсутствует, но это отсутствие лишь кажущееся. Нога Cephalopoda сильно видоизменена, и ей соответствует сово- 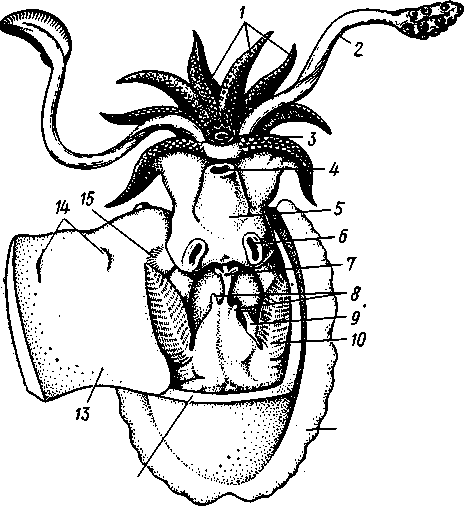 11 12 487 Рис. 482. Каракатица Sepia officinalis со вскрытой мантийной полостью, вид с брюшной стороны (по Пфуртшеллеру): 1 — руки с присосками, 2 — ловчая рука, 3 — рот, 4 — отверстие воронки, 5 — воронка, 6 — хрящевые ямки запонок, 7 — анальный сосочек с анальным отверстием, 8 — почечные сосочки, 9 — непарный половой сосочек, 10 — жабры, //— плавник, 12 — линия отреза мантии, 13 — отогнутая мантия, 14 — хрящевые бугорки запонок. /5 — мантийный звездчатый ганглий купность двух образований: воронки и щупалец. Гомология воронки ноге доказывается ее положением на брюшной стороне туловища, а также тем, что у древнего и примитивного Nautilus она не в виде трубки, а в виде желоба, раскрытого по срединной брюшной линии, т. е. очень напоминает ногу некоторых плавающих Gastropoda. У зародышей всех Cephalopoda воронка двухлопастная, и лопасти ее лишь вторично срастаются своими боковыми краями в трубку. Щупальца во время развития закладываются позади рта, на брюшной стороне тела, и лишь позднее надвигаются с двух сторон на голову, образуя околоротовой венец. Следовательно, по первичному положению щупальца соответствуют передней части ноги. Самым же важным доказательством указанного происхождения воронки и щупалец является иннервация их от педальных ганглиев. 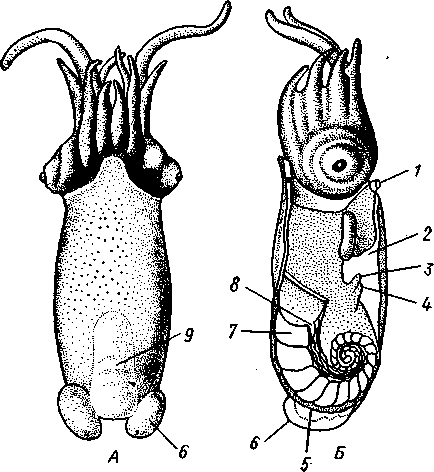 Раковина большей частью рудиментарна и хорошо развита лишь у наиболее древней из сохранившихся до сих пор форм, у кораблика Nauilus (см. рис. 481). Известковая наружная раковина наутилуса завита в одной плоскости на спинную сторону. Раковина обращена завитком вперед и замечательна тем, что полость ее разделена поперечными пере- 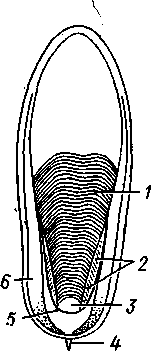 Рис. 483. Двужаберный толстоногий моллюск Spirilla со спиральной многокамерной раковиной (из Натали). А — вид со спинной стороны; Б — продольный разрез: / — воронка, 2 — мантийная полость. 3 — анальное отверстие, 4 — выделительное отверстие, S — орган свечения, 6 — плавник, 7 — раковина, 8 — сифон, 9 — участок раковины, покрытый мантией Рис. 484. Раковина каракатицы Sepia officinalis с брюшной стороны (по Мейеру) ni_. tut. галииппа лазала inij,Di иь/укл I// / i-^'n+м j ^ vypiuuinv-ri v.ivpwnui \iiiy I'ltncyy j . — сближенные перегородки спинной стороны фрагмокона. 2—боковой край проостракума, — сифональная ямка, 4 — рострум, 5 — рудимент брюшной стенки сифональной трубки, 6 — задний кпяй ппоостпакума 6 — задний край проостракума городками на ряд камер. Тело животного помещается лишь в последней, ближайшей к устью и самой большой камере (жилая камера), тогда как все прочие заполнены газом и некоторым количеством жидкости. При изменении объемных соотношений газа и жидкости животное может всплывать или погружаться на значительную глубину, используя раковину как своеобразный гидростатический аппарат. Посередине 488 каждой перегородки имеется по небольшому отверстию. Через них все камеры пронизываются тонким цилиндрическим выростом задней части туловища — сифоном. Самая первая и маленькая камера раковины называется эмбриональной. В палеозойских и мезозойских отложениях найдены остатки нескольких тысяч видов класса Cephalopoda с раковиной, построенной в общем по типу Nautilus. У современных головоногих раковина более или менее редуцируется, уменьшается в размерах и обрастает боковыми складками мантии, становясь внутренней. У интересного глубоководного рода Spirula (рис. 483) есть такая же, как у наутилуса, многокамерная раковина, но она так мала, что последняя камера ее содержит лишь небольшую часть животного и тело спирулы обрастает раковину со всех сторон. У других представителей класса Cephalopoda, еще сохранивших раковину, она в виде известковой пластинки сложного строения, залегающей под покровами на спинной стороне туловища. Такова, например, раковина каракатицы Sepia (рис. 484). Спинная сторона такой раковины прикрыта тонким роговым листком (проостракум), а задний конец образует очень маленький конус, заканчивающийся известковым рожком (рострумом). Главную массу раковинной пластинки составляет множество плотных известковых пластиночек, отходящих от проостракума косо книзу (к брюшной стороне тела). Промежутки между пластиночками очень узки и заняты рыхлой известковой массой. Происхождение такой раковины выясняется при сравнении ее с раковиной вымерших мезозойских белемнитов Belemnites (рис. 485, В).Раковина их в виде длинного прямого конуса со множеством очень узких камер, разделенных перегородками. Такая раковина называется фрагмоконом. Перегородки пронизаны сифоном, проходящим ближе к брюшной стороне конуса. Его спинная сторона выдается вперед в виде тонкого листовидного рогового продолжения. Задний конец одет мощным коническим известковым фут- 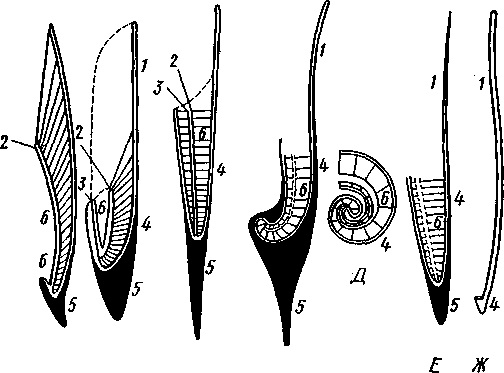 J В Г 439 Рис. 485. Схема строения раковины различных головоногих в сагиттальном разрезе; брюшная сторона раковины обращена налево. А — Sepia; Б — Belosepia; В — Belemnites (ископаемая) ; Г — Spirulirostra (ископаемая); Д — Spirula; Е — Ostra-coteuthis (ископаемая); Ж — Ommastrephes; 3 — Loligopsis (из Гешелера); / — проостракум, 2 — спинной край сифональной трубки, 3 — брюшной край сифональной трубки, 4 — совокупность камер, называемая фрагмоконом, 5 — рострум, 6 — полость сифональной трубки ляром. Обыкновенно от раковин белемнитов сохраняются только эти Концевые футляры, называемые в народе «чертовыми пальцами» и достигающие в длину 10—20 см и более. Строение раковины Belosepia — ископаемого родича каракатицы (рис. 485, Б) позволяет понять, что роговой проостракум Sepia отвечает спинной пластинке Belemnites, маленький рострум каракатицы у Belem-  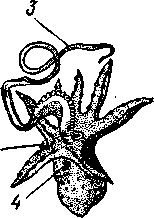 . 5 Рис. 486. Аргонавт Argonauta. A — самка, сидящая в раковине (уменьшено); Б — самец (увеличено) (из Догеля): 1 —• раковина, просвечивающая сквозь покрывающие ее расширения руки (2), 3 — гектокотиль, 4 — глаз, 5 — воронка nites был представлен громадным чертовым пальцем, а от конической раковины белемнита сохраняется, в основном, ее спинная часть, тогда как брюшная почти полностью исчезла. Перегородки спинной части фрагмокона чрезвычайно сблизились (косые пластиночки раковины Sepia) и промежутки между ними заполнились известью, так что эта часть раковины приняла вид слоистой пористой известковой пластинки. Задний конец фрагмокона сохранил) однако, не только спинную, но и брюшную стенку (рис. 485, А),образуя маленькую вороночку — сифональную ямку (см. рис. 484), полость которой представляет собой сохранившийся задний участок сифона. У некоторых других современных головоногих, например у кальмара (Lotigo), от всей раковины сохраняется лишь спинной роговой листок (проостракум), скрытый под покровами, а у большинства Octopoda раковина совершенно исчезает. Своеобразной «раковиной» обладают Argonauta из Octopoda (рис. 486). Раковина есть только у самок. Она спирально закручена, но не разделена на камеры и очень тонка. Это вторичное образование, не имеющее ничего обшего с раковиной прочих моллюсков, и выделяющееся не мантией, а эпителием особых лопастей щупалец. Раковина Argonauta служит для вынашивания яиц. Покровы. Кожа головоногих состоит из однослойного цилиндрического эпителия и подстилающего его слоя соединительной ткани. Cephalopoda обнаруживают замечательную по быстроте и резкости смену окраски, которая находится под контролем нервной системы. Смену цветов вызывает изменение формы многочисленных пигментных клеток, или хроматофоров, залегающих в соединительнотканом слое кожи. Внутренний скелет. Интересно, что кроме наружного скелета или раковины у головоногих есть еще особый внутренний скелет, служащий . 490 главным образом для защиты центральной нервной^системы. Он образован хрящом, сходным с таковым позвоночных. Хрящ в виде широкого кольца охватывает головное скопление ганглиев, образуя головную хрящевую капсулу, которая дает от себя выросты, окружающие глаза и ста-тоцисты. Кроме того, опорные хрящи развиты в запонках, в основании щупалец, внутри плавников. Как видно из описания, головная капсула в физиологическом отношении соответствует черепу позвоночных. 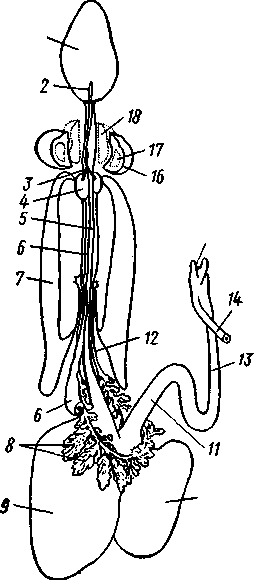 1 15 Пищеварительный канал у Cephalopoda устроен очень сложно (рис. 487). Рот лежит в центре венца щупалец и ведет в сильно развитую, мускулистую глотку. В глотке имеется язык с радулой, но терка играет в захвате и размельчении пищи второстепенную роль. Главное значение имеют две толстые роговые челюсти — спинная и брюшная, загнутые крючковидно и похожие на клюв попугая. В глотку впадают протоки одной или двух пар слюнных желез. Секрет их содержит ряд гидролитических ферментов, обеспечивающих расщепление, полисахаридов и белков. Выделения задней пары желез ядовиты. Длинный пищевод нередко образует расширение, зоб (отр. Octopoda). Пищевод переходит в мускулистый мешковидный желудок, занимающий заднюю часть пищеварительного комплекса. От той же передней стороны желудка, куда впадает пищевод, начинается тонкая кишка, которая направляется вперед. Она ведет к порошице, лежащей на особом сосочке на брюшной стороне туловища (внутри мантийной полости). 10 Рис. 487. Пищеварительная система каракатицы Sepia officinalis, вид с брюшной стороны (по Резелеру и Лампрехту): / — глотка, 2 — общий слкшной проток, 3 — слюнной проток, 4 — задняя слюнная железа, 5 — пищевод, 6 — головная аорта, 7 — печень, 8 — поджелудочная железа, 9 — желудок, 10 — слепой мешок желудка, // — тонкая кишка, 12 — печеночный проток, 13 — прямая кишка, 14 — проток чернильного мешка, 15 — анус, 16 — головная хрящевая капсула (разрезана), 17 — полость капсулы статоциста, 18 — нервное кольцо (разрезано) В желудок впадают протоки двухлопастной, реже простой печени, секрет которой содержит широкий спектр пищеварительных ферментов. Печеночные протоки большей частью усажены многочисленными небольшими железистыми придатками, выделяющими ферменты, гидролизующие полисаха-риды. Эти придатки называются поджелудочной железой. Как видно, печень и поджелудочная железа головоногих отнюдь не соответствуют в точности одноименным образованиям у позвоночных. В заднюю кишку, перед самой порошицей, открывается проток так называемого чернильного мешка. Это большая грушевидная железа, выделяющая черную, как чернила, жидкость. Несколько капель этого секрета достаточно, чтобы замутить большое количество воды. Выбрасывая жидкость через порошицу, а далее через отверстие воронки наружу, Cephalopoda окружают себя в воде темным облаком и под покровом его ускользают от преследования. Все головоногие — хищники; они нападают на различных ракообразных, а иногда и на рыб, которых схватывают щупальцами и убивают челюстями и ядом слюнных желез. 491 Нервная система головоногих достигает высокой сложности строения (рис. 488, Б).Ганглии ее велики и образуют общую окологлоточную нервную массу. Лишь на разрезах можно различить состав этой массы из нескольких ганглиев: ясно различимы парный церебральный ганглий и большой висцеральный. Что касается пары педальных ганглиев, то характерно подразделение каждого из них на два нервных узла: брахиаль-ный, или ганглий щупалец, и инфундибулярный, или ганглий воронки. От заднего отдела ганглиозной массы отходят 2 крупных мантийных нерва, которые образуют на внутренней поверхности мантии, по бокам в передней части туловища, 2 больших звездчатых гзнглия. У более примитивных Tetrabranchia (Nautilus) центральная нервная система устроена значительно проще (рис. 488, А);она состоит из трех коротких нервных дуг, одной надглоточной и двух подглоточных, причем по бокам пищевода все дуги соединяются между собой. Дуги сплошь усеяны ганг-лиозными клетками и носят характер нервных тяжей (как у Amphineura). Органами чувств головоногие снабжены богато. Для чувства обоняния служат либо осфрадии, расположенные у основания жабр (Nautilus),либо пара расположенных под глазами обонятельных ямочек (Dib-ranchia), небольших кожных углублений, выстланных чувствительным эпителием и иннервируемых от церебральных ганглиев. Острота обоняния довольно велика, это показывают опыты над ослепленными осьминогами (Octopus), которые чувствуют положенную в аквариум рыбу на .расстоянии 1,5 м. Имеется пара статоцистов, заключенных внутри хрящевой головной капсулы. Удаление их ведет к утрате способности ориентироваться в пространстве. Органы зрения Tetrabranchia могут быть сведены к типу строения крупных глазных ямок, полость которых маленьким отверстием еще со- 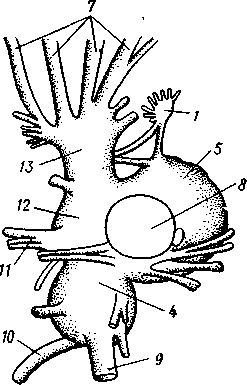 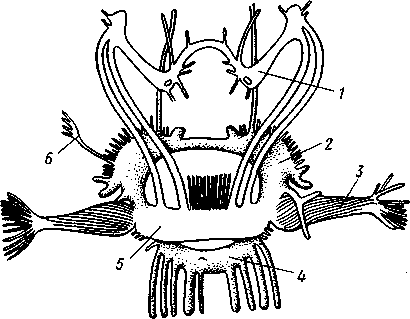 Рис. 488. Центральная нервная система головоногих. А — Tetrabranchia (Nautilus), вид спереди (из Кестнера, по Гриффину); Б — Dibranchia (Sepia), вид сбоку (из Кестнера, по Хиллигу, с изменениями): |