В.А. Догель - Зоология беспозвоночных. Тип губки (spongia, или porifera)
 Скачать 15 Mb. Скачать 15 Mb.
|

|
463 орган (пенис). Яйцевод расширяется и образует матку, в которую впадают протоки пальцевидных желез. Матка при помощи влагалища открывается в особое впячивание стенки тела — половую клоаку, куда открывается и мешок совокупительного органа. Кроме того, во влагалище впадают еще два мешковидных образования — семяприемник, предназначенный для восприятия семени другой особи, и мешок с «любовной стрелой»; стрела — это игла из углекислой извести, которая при копуля- 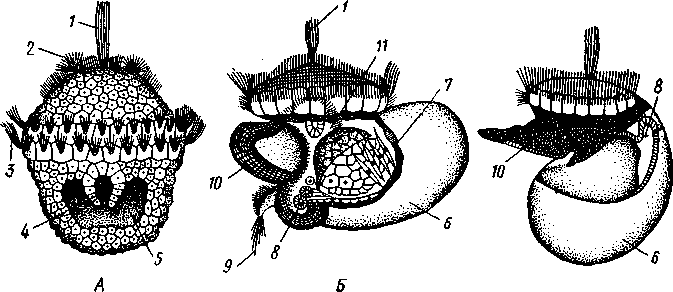 в Рис. 454. Развитие морского блюдечка Patella (Prosobranchia, Diotocardia) (по Паттен). А — трохофора; Б — парусник до скручивания; В — парусник после скручивания: 1 — теменной султан, 2 — реснички верхнего полушария, 3 — прототрох, 4 — рот, 5 — просвечивающие мезодермальные полоски, 6 — раковина, 7 — внутренностный мешок, в — мантия, 9 — реснички задней части тела, 10 — зачаток ноги, 11 — кишечник ции втыкается в кожу другой особи и служит для ее раздражения. Оплодотворение перекрестное. Развитие. Среди примитивных брюхоногих встречаются формы, обладающие наружным оплодотворением, у большинства же форм оплодотворение внутреннее. Яйца часто окружаются пергаментными или студенистыми коконами и соединяются в кладки. Дробление яйца чрезвычайно напоминает таковое Polychaeta (с. 261). Оно полное, неравномерное, детерминированное, протекает по спиральному типу. Имеются те же макромеры А, В, С и D с четырьмя квартетами микромеров. Главная часть мезодермы тоже формируется за счет клетки D — из ее потомка 4d. Целомическая мезодерма образует две мезодермальные полоски, происходящие от двух первичных мезодермальных клеток. У низших Prosobranchia из яйца развивается настоящая трохофора с предротовым венчиком ресниц (прототрохом) и с теменным султаном (рис. 454, Л). Затем трохофора превращается в личинку парусник, или велигер (рис. 454, В).У большинства представителей подкл. Prosobranchia и Opisthobranchia из яйца выходит сразу парусник. Парусник часто характеризуется развитием на предротовом отделе тела 2—4 боковых лопастей, окаймленных предротовым венчиком ресниц. Лопасти образуют мерцательный парус, от которого и сама личинка получила свое наименование. Парусник обнаруживает ряд признаков, специфичных для моллюсков. На его спинной стороне появляется утолщение эктодермы, которое затем углубляется и дает легкое впячивание — раковинную железу. На дне впячивания выделяется нежный роговой зачаток раковины; впячивание затем выворачивается, зачаток раковины занимает поверхностное 464 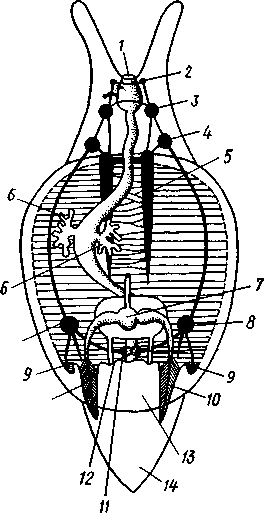 8 положение и разрастается. На брюшной стороне тела, позади рта личинки, образуется выступ, нога, на которой в виде двух маленьких боковых впячивании эктодермы закладываются статоцисты. Впереди рта появляются щупальца и глаза. Позади зачатка раковины эктодерма образует складку, нависающую над порошицей и прикрывающую зачаток мантийной полости. У легочных мантийная складка продолжает расти вперед и становится глубже лишь на правой стороне тела, постепенно перемещаясь к переднему концу. У низших брюхоногих вместо этого происходит быстрый поворот зачатка мантийной полости с порошицей и раковины на 180° (рис. 454, Б, В),вследствие чего порошица ложится на переднем конце туловища, позади головы. Свободноплавающий парусник испытывает в конце превращения редукцию предротового венчика ресниц, опускается на дно и переходит к ползающему образу жизни. Описанный метаморфоз характерен для большинства представителей подклассов Prosobranchia и Opisthobran-chia; у Pulmonata развитие прямое и протекает до конца внутри яйцевой оболочки. 10 Происхождение асимметрии. Ряд ученых старались разгадать происхождение асимметрии у брюхоногих, но часть высказанных гипотез ныне имеет лишь исторический интерес. Рис. 455. Гипотетический симметричный предок Gastropoda (по Штемпелю): 7 — рот, 3 — буккальный ганглий, 3 — церебральный ганглий, 4 — плевральные ганглии, 5 — педальный ствол, 6 — печень, 7 — желудочек сердца, 8 — париетальные ганглии, 9 — осфрадии, 10 — ктенидии, // — висцеральные ганглии, 12 — отверстие почки, 13 — мантийная полость, 14 — задний конец ноги Наиболее современной и обоснованной является теория Нэфа (1913). Есть все основания предполагать, что предками класса Gastropoda были вполне симметричные моллюски с порошицей и мантийным комплексом на задней стороне тела (рис.455), но имели раковину, закрученную в одной плоскости. Такая раковина нисколько не нарушала билатеральной симметрии. Так как вход в мантийную полость, а следовательно, и устье раковины находились сзади, то завиток раковины помещался впереди, над головой. Такая раковина несвойственна современным брюхоногим, но она характерна для примитивного головоногого моллюска — кораблика (Nautilus, с. 486). Кораблик ведет плавающий образ жизни, его раковина с завитком, обращенным вперед и закрученным в одной плоскости, не препятствует плаванию. При ползании по дну такая раковина, напротив, была бы значительной помехой, завиток ее давил бы на голову, а край устья скреб бы дно. Поэтому естественно допустить, что предок брюхоногих вел плавающий образ жизни. Причиной появления характерных черт организации Gastropoda, по Нэфу, послужил переход их предка от плавающего к ползающему образу жизни. При этом сразу же проявились отмеченные неудобства рако-кины с завитком, обращенным вперед. Устранение этих неудобств происходило следующим образом. Характерной особенностью организации брюхоногих и их ближайших предков является разделение тела на 2 области — голову с ногой и внутренностный мешок, связанные узким 465 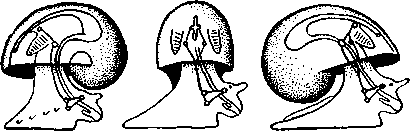 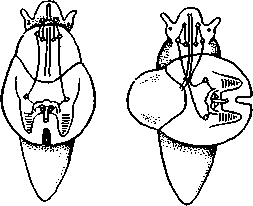 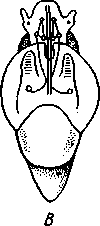 мускулистым стебельком — основанием внутренностного мешка. Относительное положение главных частей тела может значительно изменяться благодаря мышечным сокращениям стебелька. Это прекрасно видно на современных брюхоногих, способных поворачивать раковину с лежащим внутри нее внутренностным мешком. При этом голова и тесно связанная с ней нога сохраняют свое нормальное положение, а внутренностный мешок может мышечными усилиями поворачиваться вокруг вертикальной оси на 180° и более. Такие временные повороты внутренностного мешка на стебельке называются физиологической тор-сией (скручиванием). Рис. 456. Схема происхождения переднего положения мантийного комплекса и хиастонсв-рии. Верхний ряд — вид сбоку, нижний — со спинной стороны (по Нэфу). А—исходная форма со спиральной симметричной раковиной, обращенной вершиной вперед; Б — промежуточная стадия; В — форма с законченным скручиванием Перейдя к ползанию по дну, предок Gastropoda вынужден был повертывать внутренностный мешок с раковиной (рис. 456). Поворот на 180° оказался наиболее выгодным. Сначала он носил характер временной физиологической торсии, но потом новое положение раковины — завитком назад, а устьем вперед — стало постоянным и закрепилось наследственно. Именно такое положение раковины и характерно для современных представителей кл. Gastropoda. Замечательно, что молодой парусник низших переднежаберных имеет сначала раковину, обращенную завитком вперед (см. рис. 454,5), мантийные органы развиваются у него на задней стороне тела. Затем, опустившись на дно и переходя к ползанию, он испытывает торсионный процесс — вследствие сокращения мышц раковина с внутренностным мешком сразу (иногда в течение нескольких минут) поворачивается на 180° вершиной назад (см. рис. 454,В).У высших форм класса Prosobran-chia этот процесс происходит на более ранних стадиях развития и сводится к неравномерному росту: левая сторона основания внутренност-  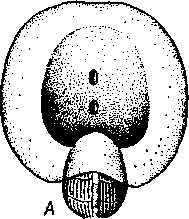 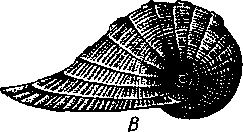 Б — Рис. 457. Раковина Tremanolus (Bellerophontidae) (из Мура). А — снизу; сверху; В — сбоку 466 ного мешка растет быстрее правой. Поэтому у них из яйца выходит парусник с раковиной, обращенной вершиной назад и с передним положением мантийного комплекса. Итак, у первичных древних брюхоногих раковина повернулась на 180°. Мантийные органы при этом оказались над головой на передней стороне туловища, а длинные плсвровисцеральные коннективы неизбеж- 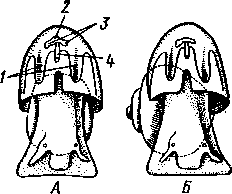 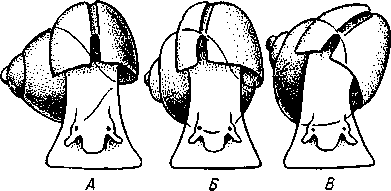 Рис. 458. Схема перехода к асимметричной па-ковине. Вид спереди (по Нэфу). А и Б — последовательные стадии: / — ктенидии, 2 — желудочек сердца, 3 — предсердия. 4 — задняя кишка Рис. 459. Схема регуляции положения раковины. Вид спереди (по Нэфу). А — первичное состояние, ось раковины поперечна; Б — ось раковины занимает косое положение, вершина сдвинута назад; В — нормальное положение раковины, вершина приподнята кверху но перекрестились, т. е. возникла хиастоиеврия (рис. 456). Раковина, повернувшаяся вершиной назад, сперва оставалась вполне симметричной. Именно такими брюхоногими, по-видимому, были ископаемые Bel-lerophontidae из нижнего палеозоя (рис. 457). Следовательно, раковина, повернутая вершиной назад, переднее положение мантийного комплекса и хиастоневрия возникли одновременно как результат единого процесса филогенетической торсии. 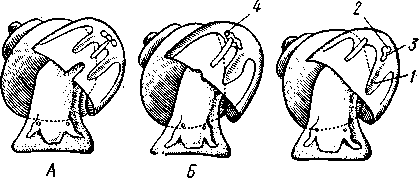 В Дальнейшие эволюционные изменения в пределах класса Gastropoda сводятся к следующему. В результате ползающего образа жизни увеличилась нога и соответственно должен был возрасти объем раковины, в которую при опасности втягиваются как голова, так и нога. Увеличение объема раковины при сохранении прежнего ее диаметра возможно лишь за счет расположения ее оборотов в разных плоскостях. Поэтому раковина приняла коническую форму, а вершина ее все более и более стала выдаваться вправо (рис. 458). Но такая асимметричная (правозакручен-ная) раковина имеет неустойчивое положение, перетягивая животное на правый бок. По-этому для сохранения равновесия онг все более отклоняется вершиной назад и вверх, а устье ее повертывается несколько вправо (рис. 459), г. е. происходит некоторое раскручивание, или деторсия. Это яв- „ ... „ Рис. 460. Схема образования асимметрии ман- ление, называемое регуляцией ТИЙНОГо комплекса. Вид спереди (по Нэфу). положения раковины, приво- А — В — стадии постепенной редукции правого дит к следующему: 1. Внут- ктенидия и предсердия и смещения задней ревностный мешок давит своей кишки направо: пприл;М7Тттрртприил i-n / —ктенидии, 2 — желудочек сердца, 3 — левое пред- Преимущественно ha сердне, < — рудимент правого предсердия 467 правую сторону мантийной полости. 2. Недостаток места в правой части мантийной полости и повышенное давление вызывают постепенную редукцию и, наконец, полную утрату правых мантийных органов (рис. 460). Так возникают характерные черты Prosobranchia Monoto-cardia (рис. 461), которые лишены правых ктенидиев, осфрадия, предсердия и почки. 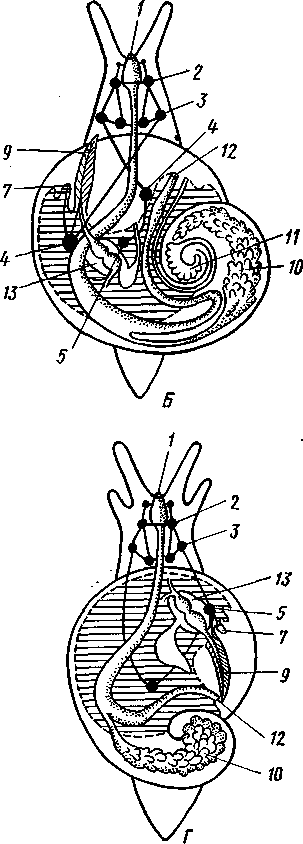 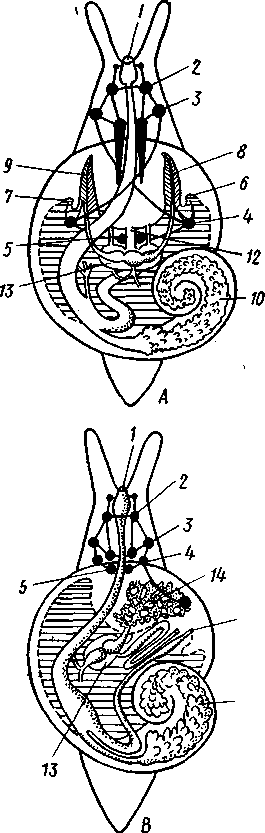 12 10 468 Рис. 461. Схема типов асимметрии у разных Gastropoda (по Штемпелю). А — Prosobranchia Diotocardia; Б — Prosobranchia Monotocardia; В — Pulmonata; Г — Opisthobranchia: / — рот, 2 — церебральный ганглий, 3—плевральный ганглий, 4—париетальный ганглий, 5 — висцеральный ганглий, 6. 7 — осфрадии, 8, 9 — ктеиидии, 10 — печень, // — половая железа. 12 — анус, 13 — перикардий, 14 — сосудистая сеть легкого При сильной деторсии (у подкл. Opisthobranchia; рис. 461.Г) мантийный комплекс оказывается на правой стороне тела, а нервная система раскручивается, вновь становясь эвтиневральной. У легочных (подкл. Pulmonata; рис. 461,5) деторсия незначительна, и мантийные органы остаются в передней части тела, их нервная система также эвтиневраль-на, но эта особенность вызвана не деторсией, а сильной концентрацией ганглиев в области глотки. Позднее Гарстранг (1928) внес существенное исправление в теорию Нэфа. По его мнению, торсионный поворот внутренностного мешка у предков Gastropoda происходил не во взрослом состоянии, а еще у планктонных личинок, как это наблюдается у низших современных брюхоногих (например, у Patella; рис. 462,Г). Этот процесс был полезен, так как только обращенная устьем вперед раковина давала возможность личинке втягивать в нее сначала более жизненно важную часть тела — голову, а потом уже ногу. Итак, это исправление Гарстранга касается только самого первого этапа становления асимметрии, все остальные и более поздние ее проявления, по-видимому, были правильно объяснены Нэфом. Экология. Большинство Prosobranchia и все Opisthobranchia — морские животные. Некоторые переднежаберные приспособились к жизни в пресной воде и даже на суше (Viviparus, Bithynia, Cyclophorus, Acme, Helicina и др.)- В подклассе Pulmonata представлены в основном сухо- 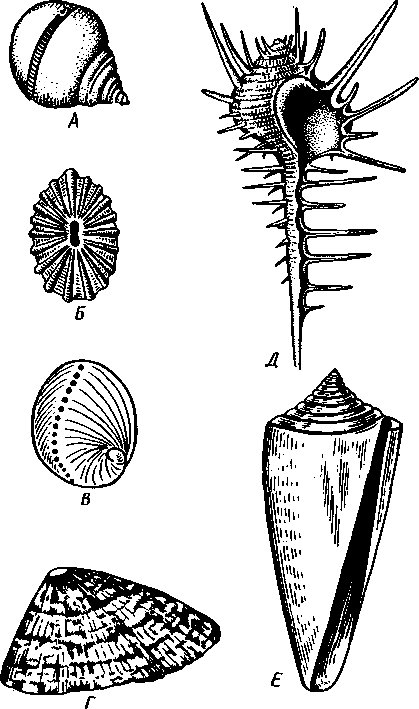 Рис. 462. Ракови ны переднежа- берных моллюс ков. А — Pleuroto- maria; Б — Fissu- rella; В — Halio- tis; Г— Patella; Д — Murex; E — Conus (A — В — по Лангу, Г — по Иванову, Д — Е — из Гаймен) 469 путные и пресноводные формы. В море брюхоногие встречаются на различных глубинах, на суше — в самых различных климатах — выносливы по отношению к температуре. Перенесение резких колебаний температур облегчается способностью многих легочных к спячке — зимней на севере, летней и зимней на юге. При этом улитка забирается в почву, втягивается в раковину и заклеивает устье эпифрагмой, т. е. пленкой из застывшего слоя слизи с большим содержанием солей извести. Переход от ползания по дну водоема к плаванию совершался у брюхоногих в различных группах независимо. Пелагическими брюхоногими являются киленогие (см. рис. 439) —Heteropoda (из подкл. Prosobranchia) и крылоногие (см. рис. 440)—Pteropoda (из подкл. Opisthobranchia); и те и другие хорошо приспособлены к плаванию в толще воды. Тело их обычно стекловидно, прозрачно, нога образует непарный килевидный плавник (Heteropoda) или превращена в пару больших крыловидных плавников (Pteropoda); раковина более или менее редуцируется, вплоть до полного исчезновения. Настоящие паразиты имеются только среди переднежаберных брюхоногих. Большинство их паразитирует на коже или в полости тела иглокожих (морские звезды, морские ежи, голотурии). Паразитический об раз жизни вызвал у этих брюхоногих (Entocolax, Parenteroxenos) сильные изменения и упрощения организации (утрата раковины, мантии, ноги и т. д. вплоть до полной редукции пищеварительной, кровеносной, нервной систем). Пища брюхоногих разнообразна. Наряду с растительноядными формами, широко распространенными в воде и на суше, имеется большое количество хищников, питающихся червями, раками или другими моллюсками. Полезное и вредное значение брюхоногих. Практическое значение брюхоногих по сравнению с другими моллюсками (пластинчатожаберными и особенно головоногими) не очень велико. В ряде европейских стран употребляется в пищу виноградная улитка (Helix pomatia), которую для этой цели разводят в специальных улитковых хозяйствах. Съедобны и многие морские переднежаберные: трубач Buccinum undaturn, береговая улитка Littorina и некоторые другие. Перламутр раковины некоторых морских Prosobranchia, например Turbo, Trochus и Haliolis, служит для выделки различных украшений, безделушек и пуговиц. Различные морские раковины, главным образом каури (Monetaria moneta), вплоть до начала XX в. служили у некоторых народов в качестве разменной монеты. Сведения о подобном использовании каури имеются, например, в летописях Индии от VII в. н. э. В XIX в. каури ввозились в большом количестве в Западную Африку для сбыта туземцам, делавшим из нанизанных в, виде бус раковин денежную единицу. Вредное значение для сельского хозяйства имеют некоторые наземные формы из подкласса Pulmonata, а именно слизни, или слизняки. Примером слизней может служить достигающий 3—6 см длины полевой слизень (Deroceras reticulatus), наносящий вред озимым посевам, а также картофелю, свекле, табаку, клеверу и огородным растениям. В южных районах значительный вред могут наносить моллюски рода Parmacella, относящиеся к группе слизней. Многие из них представляют серьезную угрозу для садов и огородов. Велико отрицательное значение брюхоногих как первых промежуточных хозяев трематод (с. 168), среди которых имеется много возбудителей опасных заболеваний человека и животных. |