В.А. Догель - Зоология беспозвоночных. Тип губки (spongia, или porifera)
 Скачать 15 Mb. Скачать 15 Mb.
|

|
КЛАСС ПОГОНОФОРЫ (POGONOPHORA) Погонофоры — своеобразные морские животные, обитающие преимущественно на больших глубинах (3—10 тыс. м) и ведущие сидячий образ жизни. Они живут в очень длинной (от 15 см до 1,5 м), тонкой и прочной защитной трубочке, состоящей из хитина, выделяемого кожными железами животного. Верхний конец трубочки выдается над илистым дном, из него может выставляться передний конец животного, снабженный щупальцами.  Рис. 560. Погонофора Choanophorus indicus (по Буб- ко). А — общий вид самца; Б — задний щетинконос- ный отдел тела: 1 — щупальца, 2 — головная лопасть, 3 — первый сегмент тела, 4 — второй сегмент тела, 5 — третий сегмент тела, 6 — папнллы, 7 — задний щетинконосный отдел тела, 8 — щетинки на заднем отделе тела 569 Большинство погонофор имеет длинное и очень тонкое тело, ширина которого нередко измеряется долями миллиметра. Гигантом среди погонофор можно назвать Lamellibrachia barhami, достигающую 0,5 м в длину при ширине тела в б мм. Строение и физиология. Внешняя морфология. Очень сильно вытянутое, почти нитевидное цилиндрическое тело погонофор (рис. 560) состоит из четырех первичных сегментов (отделов): очень короткого первого, более длинного второго, чрезвычайно вытянутого третьего, или туловища, и четвертого. Первый отдел снабжен небольшой головной лопастью и венцом щупалец. Средний отдел несет два косых валика с кутикулярными килями — уздечку. На туловище имеются многочисленные хитиноидные пластинки, лежащие на небольших возвышениях — папиллах и служащие для опоры о стенку трубки, когда животное перемещается в ней. Расположение пластинок в общем беспорядочное, но в передней и задней частях туловища они более или менее метамерны. В средней части, кроме того, развиты два пояска из мелких зубчатых пластинок, позволяющих животному прочно закрепляться в трубке. Наконец, самый задний отдел тела несет крошечные щетинки; у некоторых видов щетинки располагаются беспорядочно, у других — поперечными рядами по 4—6 в каждом ряду. Эти щетинки напоминают таковые полихет; они служат для рытья в грунте (см. ниже). Стенка тела погонофор состоит из однослойного, богатого одноклеточными железами кожного эпителия, покрытого почти сплошь кутикулой. Под ним располагается хорошо развитый кожно-мускульный мешок, состоящий из наружного тонкого слоя кольцевых мускульных волокон и внутреннего, обычно очень мощного, слоя продольных мышц. Затем следует перитонеальный эпителий, ограничивающий полость тела. Целом. Каждый сегмент тела содержит свой участок целома: в первом сегменте лежит непарный передний целом, в двух следующих сегментах— по паре целомических мешков (см. рис. 563). В последнем четвертом сегменте целом закладывается сначала в виде пары мешочков, которые позднее сливаются, образуя единую полость (см. рис. 564). У погонофор существует и зачаточное вторичное его расчленение. Во-первых, от общего целома в передней части туловища отделяются небольшие участки, находящиеся в метамерных спинных прикрепительных папиллах. Во-вторых, в заднем щетинконосном отделе тела образуются поперечные перегородки, врастающие от стенок тела в уже существующий здесь общий целом, который разделяется благодаря этому на ряд метамерных камер. Пищеварение. Замечательной особенностью погонофор является полное отсутствие кишечника. Функции собирания пищи, переваривания и всасывания выполняет аппарат щупалец. Щупальца представляют очень длинные выросты первого сегмента, внутрь которых продолжаются каналы его целома (рис. 560, 561). Наиболее примитивные формы (Oligobrachia dogieli) имеют 6—9 щупалец, расположенных на подковообразном основании (рис. 562), у других .погонофор их значительно больше (от 12 до 2000). У Siboglinum одно, но очень длинное щупальце закручивается штопорообразно, причем так плотно, что прилегающие друг к другу обороты спирали образуют полый цилиндр. Прилегая друг к другу (иногда даже спаиваясь), щупальца ограничивают межщупальцевое пространство. Вдоль каждого щупальца расположены ряды многочисленных очень тонких выростов — пиннул, торчащих в межщупальцевое пространство и переплетающихся там в виде густой сети. Мерцанием ресничек у основания пиннул через меж- 570 щупальцевое пространство прогоняется вода, причем пищевые частицы (мелкие планктонные организмы и органический детрит) застревают среди пиннул. Здесь же, в межщупальцевом пространстве, происходит переваривание пищи и всасывание питательных веществ посредством пиннул. Каждая пиннула представляет одну сильно вытянувшуюся эпи-  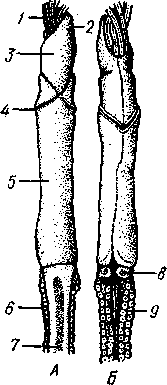 Рис. 561. Spirobrachia beklemischevi со спинной стороны (по А. Иванову): 1 — щупальца, 2 — первый и второй сегменты, 3 — уздечка, кутикулярные кили, 4 — прикрепительные органы, 5 — туловище (третий сегмент), 6 — аояски зубчатых щетинок, 7 — задняя часть туловища Рис. 562. Передний конец тела Oligobrachia dogieli с брюшной (Л) и спинной сторон (Б) (по А. Иванову): / — основание щупалец, 2 — головная лопасть, 3 — первый сегмент, 4 — уздечка, 5 — второй сегмент, S — третий сегмент (туловище), 7— спинная ресничная полоска. 8 — мужское половое отверстие, 9 — просвечивающие железы, выделяющие трубку телиальную клетку, содержит одно ядро и длинную петлю тончайшего капиллярного сосуда. Кровь попадает в этот капилляр из приносящего сосуда щупальца, а выносится из него в выносящий щупальцевый сосуд. Таким образом, межщупальцевое пространство играет роль своеобразной «кишечной полости», пиннулы — «кишечных ворсинок», а сами по-гонофоры представляют совершенно исключительный пример свободно-живущих животных, у которых обычное кишечное пищеварение полностью заменено наружным пищеварением. Кровеносная система замкнутая, состоит в основном из спинного и брюшного сосудов (рис. 563). Спинной сосуд способен сокращаться и гонит кровь сзади наперед; у основания щупалец он расширяется, образуя мускулистое сердце, к которому иногда прилегает эпителиальный пузырек перикардия, напоминающий таковой гемихордовых. Приносящие сосуды щупалец берут начало от спинного сосуда, брюшной сосуд образуется слиянием выносящих щупальцевых сосудов. Кровь окрашена в красный цвет растворенным в ней гемоглобином. Органы выделения представлены парой сильно развитых почек, открывающихся внутренними концами в передний непарный целом; они 571 представляют собой целомодукты 1-го сегмента (рис. 563). В отличие от гемихордовых 2-й сегмент погонофор без целомодуктов. В 3-м, туловищном, сегменте имеется своя пара целомодуктов, но здесь они играют роль половых протоков. 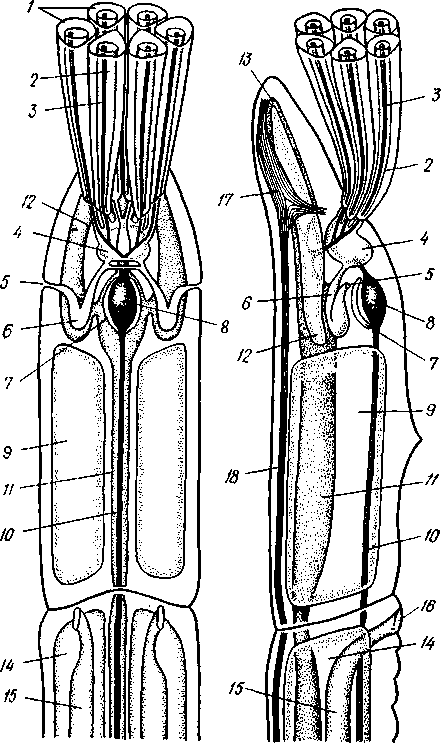 Нервная система очень примитивна и целиком залегает в толще кожного эпителия, напоминая нервную систему Hemichordata. В эпителии головной лопасти находится мозг — брюшное скопление ганглиозных клеток. От мозга назад тянется брюшной нервный тяж и берут начало нервы щупалец. На брюхе туловищного отдела имеется полоска мерцательного эпителия; возможно, это орган химического чувства. Половая система. Пого-нофоры раздельнополы, половой диморфизм отсутствует. Половая система лежит в 3-м сегменте. У самцов в задней половине туловища помещается пара длинных колбасовидных семенных мешков, открывающихся посредством длинных целомодуктов — семяпрово-водов на передней границе туловища. В семяпроводах формируются мешковидные сперматофоры, содержащие сперматозоиды и снабженные длинной нитью. У самок пара вытянутых яичников лежит в передней части туловища, а сравнительно короткие яйцеводы открываются в средней части туловища. Развитие. Самка откладывает яйца в передний участок собственной трубки, где протекает все развитие. Яйца более или менее удлиненные, богатые желтком. Рис. 563. Схема строения Pogonophora (по А. Ива-нону) . А — передний участок тела самца со спин-нон стороны; Б — то же, с левой стороны: 1 —• щупальца, 2 — цсломический канал щупальца, ,? — приносящий и уносящий сосуды щупальца, 4 — целом первого сегмента, 5 — наружное отверстие целомодукта, 6 — целомодукт первого сегмента, 7 — перикардий, 8 — сердце, 9 -- целом второго сегмента, 10 — спинной сосуд, 11 — брюшной сосуд, 12 — боковой головной сосуд, 13 — срединный головной сосуд, 14 — целом третьего сегмента, 15—семяпровод, 16 — половое отверстие, 17 — мозг, 18 — брюшной нервный ствол 572 Дробление яйца напоминает спиральное, детерминированное. Однако в отличие от кольчатых червей и моллюсков у погонофор целоми-ческая мезодерма образуется не из телобластов 4d, a за счет потомков переднего макромера В. В результате дробления образуется плотный зародыш. Мелкие клетки эктодермы постепенно обрастают крупные энтодер-мальные бластомеры, остав- 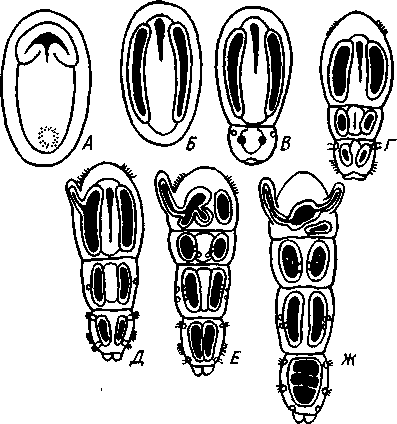 ляя на будущей брюшной стороне небольшое отверстие бластопора. В зачатке энтодермы появляется полость, и он превращается в замкнутый со всех сторон первичный кишечник. От последнего спереди отделяется пара мезодермальных мешков — зачатки целома, которые быстро растут в заднем направлении. После этого тело зародыша расчленяется на отделы, причем сначала обособляется четвертый, самый задний первичный сегмент, или отдел тела, и в него попадают задние участки целома (рис. 564). Затем формируется третий сегмент с его целомически-ми мешками, наконец, появляется граница между целомами первого и второго сегментов. Сначала развивается одно левое щупальце, в него заходит продолжение левого переднего целомического мешочка, потом одно за другим вырастают остальные щупальца. Все они сообщаются с левым передним целомом. Околосердечная сумка (перикардий) образуется из правого переднего целомического мешочка. Итак, целом первичных сег- рис. 564. Схема развития целома и расчлене- ментов у Pogonophora образу- ние тела у погонофор (по А. В. Иванову). А — ется так же как И у ИГЛОКОЖИХ появление энтероцельных выпячиваний кишеч- и китпрчнп'пытпятпих (с ^W ника; 5 —Р°ст энтероцельных целомических И кишечнодышащих (С.ООЭ), мешков; S — отделение задней пары целомиче- т. е. знтероцельным путем, ских мешков и заднего сегмента (телосомы); Внутренняя масса энтодер- Г —отделение от передних целомов метасомы; мальных клеток, оставшихся Д — развитие 1-го щупальца и появление ще-после отделения целома, пред- тинок на телосоме; Е-закладка 2-го щупаль- „ г ца, отделение от передних целомов мезоцелеи; ставляет зачаток средней киш- ж-образование септ в задних целомах (те ки, однако далее ОН не разви- лосоме) и формирование непарного переднего вается и в конце эмбриональ- целома (протоцеля) ного развития рассасывается. В дальнейшем на переднем и заднем концах зародыша развиваются ресничные пояски, но они временные и у взрослого животного не сохраняются. Молодое животное выходит из материнской трубки и сразу же переходит к жизни на дне, выделяя собственную трубку. Дальнейшее развитие сводится к усиленному росту, образованию прикрепительных органов, половых органов и т. д. Свободноплавающая личинка в развитии изученных до сих пор в эмбриологическом отношении погонофор отсутствует. Филогения. Образ жизни погонофор сходен с таковым сидячих поли-хет. Не удивительно поэтому, что и внешний вид, и даже некоторые детали наружного строения в этих двух группах обнаруживают много общего. Конвергентное поверхностное сходство послужило некоторым зоологам поводом неправильно рассматривать погонофор как аберрантных многощетинковых кольчецов из подкласса Sedentaria. В организации погонофор мы видим много характерных особенностей, присущих также аннелидам, эхиуридам, моллюскам и отчасти щупальцевым и ше-• тинкочелюстным. Это, с одной стороны, способность образовывать хитин 573 и щетинки «аннелидного типа». С другой стороны, погонофоры сходны с частью щупальцевых (плеченогими), с щетинкочелюстными и с вто-ричноротыми энтероцельным способом образования вторичной полости тела, а также асимметрическим развитием передней пары целомиче-ских мешков, образованием перикардия из правого переднего целома. Подобно аннелидам и хордовым, погонофоры — животные сегментированные и полимерные (состоящие из многих сегментов). Однако сегментация в этих трех группах развилась независимо и различными путями. У погонофор наиболее отчетливо сегментирован задний отдел тела, и это объясняется особенностями их образа жизни. Дело в том, что погонофора в течение жизни почти непрерывно закапывается в грунт задним концом, который приспособлен для активного рытья и выдвигается для этого через заднее отверстие трубки. Метамерные щетинки и мускулистые поперечные перегородки, разделяющие целом заднего отдела тела, развились как необходимые приспособления к активному рытью. По мере зарывания животного в ил трубка сзади постепенно удлиняется. Вероятно, эволюция погонофор пошла по пути глубокого приспособления к сидячему образу жизни в защитной трубке, причем одним из своеобразных процессов явилась редукция кишечника и физиологическая замена его щупальцевым аппаратом. Эволюция погонофор сопровождалась множественной закладкой прикрепительных хитиноидных пластиночек и вторичным упорядочиванием их расположения. Щупальцевый аппарат в одних случаях испытал оли-гомеризацию (единственное щупальце Siboglinum), в других, напротив, количество щупалец значительно возросло (Lamellisabella, Spirobrachia и Lamellibrachia). ТИП ЩЕТИНКОЧЕЛЮСТНЫЕ (CHAETOGNATHA) Щетинкочелюстные — небольшая, своеобразная, довольно обособленная от других типов группа морских планктонных животных. Наиболее вероятно их родство с Deuterostomia, на что особенно указывает их эмбриональное развитие. 1. Тело щетинкочелюстных состоит из трех отделов — головы, туловища и хвоста, причем каждый отдел имеет собственный участок целома. 2. На голове расположены две боковые группы мощных крючкооб-разных хватательных щетинок. 3. Тело окаймлено плавниками — парными боковыми и хвостовым. 4. Нервная система состоит из надглоточного и подглоточного ганглиев, соединенных окологлоточными коннективами, и подкожного нервного сплетения. 5. Кровеносная и выделительная системы отсутствуют. 6. Во время эмбрионального развития целом образуется энтероцельным путем. К типу Chaetognatha принадлежит только один класс с тем же названием. КЛАСС ЩЕТИНКОЧЕЛЮСТНЫЕ (CHAETOGNATHA) Известно всего 50 видов щетинкочелюстных, большая часть которых в течение всей жизни обитает в планктоне. Однако представители рода Spadella ведут донный образ жизни. Щетинкочелюстные— быстрые, плавающие как бы толчками (вследствие характерных изгибаний тела в спиннобрюшном направлении) хищники, поедающие планктонных 574 животных и нападающие порой на добычу, превосходящую их размерами. Тело щетинкочелюстных стекловиднопрозрачно; по наличию плавников и по характеру движения они несколько напоминают маленьких рыбок. Размеры тела колеблются от 1 до нескольких сантиметров, Sagiita gazellae имеет 10 см в длину. Строение. Удлиненное тело несколько сужается к концам (рис. 564). На заднем конце и по бокам тела кожа образует выдающиеся складки: хвостовой и боковые плавники. Тело явственно делится двумя поперечными перегородками на три отдела: головной, туловищный и хвостовой. Головной конец прикрыт сверху и с боков двойной кожной складкой — капюшоном. Тело покрыто многослойным кожным эпителием (редкий случай среди беспозвоночных), под которым имеется слой продольных мышц, имеющих поперечнополосатое строение. Мускулатура подстилается изнутри перитонеальным эпителием, ограничивающим обширный целом. Кишечник в виде прямой трубки; начинается ртом на переднем конце тела и кончается порошицей на границе между туловищем и хвостом. Рот окружен с боков двумя рядами хитиновых подвижных крючков, отчасти прикрытых капюшоном. Кишечник подвешен на спинно-брюш-ном мезентерии, делящем полость тела на правую и левую половины. Chaetognatha — хищники. Нервная система состоит из надглоточного и подглоточного ганглиев, образующих мозг, и из мощного брюшного узла, лежащего в туловище под кишечником. Мозг и брюшной узел соединены длинными коннек-тивами; брюшной ганглий дает 12 пар нервов к стенке тела, где они соединяются с хорошо развитым подкожным нервным сплетением. Над мозгом лежит пара маленьких глаз, причем каждый из них состоит из трех слившихся глазков, о чем можно судить по наличию трех хрусталиков. Глаза инвертированы. Позади глаз, на затылке, имеется площадка мерцательного эпителия, которой придают значение органа обоняния. Кроме того, по телу рассеяно значительное количество осязательных эпителиальных бугорков, усаженных на вершине чувствительными волосками. Половая система. Chaetognatha — гермафродиты. Женские половые органы помещаются в задней части туловища, мужские — в хвостовом отделе. По бокам задней кишки лежат два яичника, от них отходят яйцеводы, открывающиеся наружу на боковых стенках тела. Интересно, что внутренние, обращенные к яичникам концы яйцеводов замкнуты слепо, так что сообщение с яичниками устанавливается, вероятно, лишь в период половой деятельности. В каждом из боковых отделов хвостовой полости тела имеется по массивному клеточному валику, семеннику, прилегающему к стенке тела. Созревающие живчики попадают в целом и выводятся наружу через два семяпровода, которые начинаются широкой воронкой и открываются наружу по бокам хвостового отдела. Каждый семяпровод образует на своем пути расширение — семенной пузырь, в котором скапливаются живчики. Размножение исключительно половое. |