В. Н. Сайтаниди Рецензент членкорреспондент расхн в. Ф. Красота Петухов В. Л. и др. П31 Ветеринарная генетика В. Л. Петухов, А. И. Жигачев, Г. А. Назарова. 2е изд., перераб и доп. М. Колос, 1996. 384 с ил. Учебники
 Скачать 5.3 Mb. Скачать 5.3 Mb.
|

|
НАСЛЕДОВАНИЕ ГРУПП КРОВИ У всех видов животных большинство аллелей генетических систем групп крови наследуется по типу кодоминирования, т. е. в гетерозиготе фенотипически проявляются оба гена. Весьма редко встречаются рецессивные аллели, подобные аллелю О системы АВО у человека. В связи с этим возможен анализ частоты аллелей разных локусов в популяциях во времени и в пространстве, что является главным инструментом для описания их генетической структуры и позволяет приблизиться к пониманию эволюционного процесса. Все известные системы групп крови у сельскохозяйственных животных локализованы в аутосомах. В сложных системах (у скота В- и С-системы) антигенные факторы контролируются несколькими тесно сцепленными сублокусами. С-система состо- 212 ит из двух серий аллельных (или почти близко к аллельным) генетических детерминант Сь С2, С'{ ,С2 и Хь Х2) С, Fi0. Анализ рекомбинаций между концевыми антигенами С-системы показал, что длина участка ДНК этой системы составляет 0,3 сМ (сантиморгана), тогда как В-системы — 0,7 сМ. Можно выделить три основных правила наследования групп крови: 1) каждая особь наследует по одному из двух аллелей от отца и от матери в каждой системе групп крови; 2) особь с антигенами, не обнаруженными хотя бы у одного из родителей S^^SST быть потомком данной родительской пары (например, Р $ Fp/F х d'FF/V * FiN/N); 3) гомозиготная особь по одному антигену, например F/F, не может быть потомком гомозиготной особи с противоположным антигеном, например V/V. Получение реагентов для определения групп крови. Иммунологическая специфичность белковых антигенов определяется: 1) последовательностью аминокислот полипептидной цепи (иногда изменение даже нескольких аминокислот ведет к изменению антигена); 2) концевыми аминокислотами цепи; 3) вторичной структурой белковой молекулы; 4) наиболее активными поверхностно расположенными участками полипептидной цепи — антигенными детерминантами (один антиген может иметь несколько детерминантных групп). Антигены выявляются при помощи реакции антиген — антитело. Основой для определения взаимодействия антиген — антитело служит у крупного рогатого скота и овец реакция гемолиза (разрушение стромы эритроцитов с выделением из них гемоглобина), у свиней — полная и неполная агглютинация (склеивание эритроцитов) и реакция гемолиза. Схема получения моноспецифической сыворотки В приведена на рисунке 42. Кровь от животного-донора, имеющего антигены Ас, Ва и Са, вводят реципиенту с антигеном Ас, но не имеющему антигенов Ва и Са. У реципиента вырабатываются антитела к антигенным факторам Ва и Са. Антитела против антигена Ас не образуются, так как у реципиента есть этот фактор. В сырой сыворотке абсорбируют ненужные антитела, в данном случае анти-Са, эритроцитами третьего животного, имеющего антиген Са. Потом из сыворотки путем центрифугирования удаляют эритроциты с абсорбированными на них антителами Са. Полученную моноспецифическую сыворотку можно использовать для выявления антигена Ва в эритроцитах других животных. Системы групп крови. В настоящее время у крупного рогатого скота открыто 12 систем групп крови, у свиней — 17, у овец — 16, у лошадей — 9, у птиц — 14. Из всех этих систем наиболее сложной является В-система у крупного рогатого скота, включающая более 40 антигенов, которые в различных комбинациях образуют более 500 аллелей. Если в системе имеется более трех 213 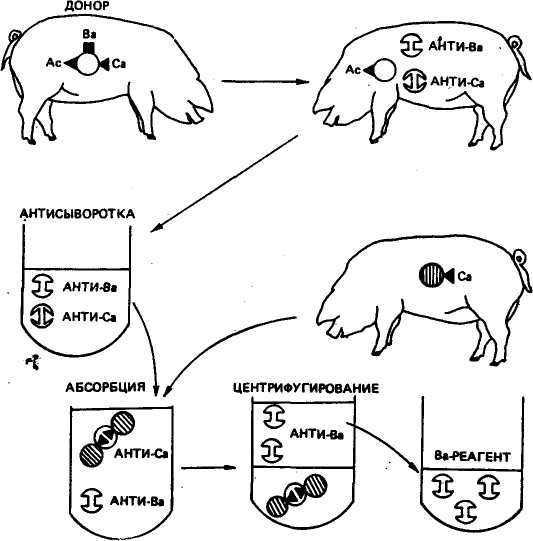 РЕЦИПИЕНТ ОБОЗНАЧЕНИЯ: ЭРИТРОЦИТЫ С АНТИГЕННЫМ ФАКТОРОМ Ас ЭРИТРОЦИТЫ С АНТИГЕННЫМ ФАКТОРОМ Ва ЭРИТРОЦИТЫ С АНТИГЕННЫМ ФАКТОРОМ Са 5£ АНТИТЕЛА ПРОТИВ АНТИГЕНА В В СЫВОРОТКЕ 5J АНТИТЕЛА ПРОТИВ АНТИГЕНА С В СЫВОРОТКЕ Рис. 42. Схема получения моноспецифической сыворотки путем иммунизации аллелей, то такие системы называют полиаллельными. К ним кроме системы В относят системы С, S, А, у свиней — Е, L, М, у овец — В, А, С. J-система крупного рогатого скота имеет иммуногенетическое сходство с антигеном А человека, свиней и антигеном R овец, S-система гомологична М-системе овец. Система Р групп крови у лошади аналогична АВО-системе человека. У крупного рогатого скота установлена связь J-системы с локусоМ| гемоглобина (НЬ) и Р-лактоглобулина (pL) ЗНАЧЕНИЕ ГРУПП КРОВИ ДЛЯ ПРАКТИКИ Контроль достоверности происхождения животных. Одна из главных областей практического применения групп крови — контроль происхождения животных. Такое их использование вызвано тем, что в некоторых стадах встречается 20 % и более ошибок в происхождении животных. Это может быть следствием не только недостатков в работе техников по искусственному осеменению, потери номеров, неправильного их чтения, но и результатом повторных осеменений животных спермой разных производителей (в повторную охоту приходит до 50 % коров, а продолжительность стельности в норме изменяется от 270 до 292 дней) и других случаев. Контроль происхождения необходим и при испытании свиноматок по качеству потомства, осемененных смешанной спермой хряков (В. Н. Тихонов, 1967), для установления моно- и дизи-готности двоен, при получении животных методом трансплантации эмбрионов и т. д. Контроль достоверности происхождения животных возможен благодаря: 1) кодоминантному наследованию антигенных факторов; 2) их неизменности в течение онтогенеза; 3) огромному числу комбинаций групп крови, которые в пределах вида практически не бывают одинаковыми у двух особей, за исключением монозиготных близнецов. В таблице 33 приведен пример уточнения отцовства в случае, когда корова в первый раз и повторно была осеменена спермой разных быков. По системе А невозможно уточнить происхождение потомка, так как аллель DH есть у обоих быков. В системе В теленок получил один аллель ВО2А от матери (такого аллеля нет у предполагаемых отцов), а второй АВ — от быка № 2 (этого аллеля нет у первого производителя). Поэтому уже можно сделать заключение, что отцом теленка является бык № 2 (исходя из второго правила). Это заключение подтверждается и наличием у потомства аллеля W в системе С. Точно так же по системе F—V можно сделать заключение, что первый производитель не может быть отцом, так как он гомозиготен по аллелю F/F, а потомок гомозиготен по противоположному аллелю V/V (третье правило). 33. Уточнение отцовства по группам крови
214 215 Иммуногенетический анализ близнецов. Как известно, близнецов, развивающихся из одной зиготы, называют монозиготными или однояйцовыми, а из двух оплодотворенных яйцеклеток (зигот) — дизиготными или двуяйцовыми. Монозиготные близнецы всегда одного пола и имеют одинаковые группы крови. Разнополые двойни всегда дизиготные и с разными группами крови. В среднем у крупного рогатого скота рождается около 2—3 % двоен, среди которых 50 % двуполых пар, 25 % пар бычков и 25 % телочек. Среди общего количества двоен только 10 % бывает монозиготных (поровну мужского и женского пола). В 90 % случаев у двоен крупного рогатого скота возникает анастомоз (срастание) кровеносных сосудов, и, как следствие этого, у дизиготных двоен наблюдается химеризм (моза-ицизм) эритроцитов. Смесь двух различных типов эритроцитов называется эритроцитарнымхимеризмом. Впервые это явление открыл Оуэн в 1945 г. у двоен крупного рогатого скота, что явилось важным вкладом в разработку теории приобретенной иммунологической толерантности. В эмбриональный период при анастомозе сосудов образуется два типа эритроцитов и антигенов, соответствующих их генотипам (рис. 43). Но в связи с обменом эритроцитов на ранней стадии онтогенеза у близнецов не образуются антитела на чужеродные ДВУЯЙЦОВЫЕ БЛИЗНЕЦЫ ПЕРВЫЙ ПЛОД ВТОРОЙ ПЛОД НАСЛЕДУЕТ НАСЛЕДУЕТ ФАКТОРА ФАКТОР В ОБА ПЛОДА НАСЛЕДУЮТ ФАКТОРЫ АИВ  ПЕРВЫЙ ПЛОД ПОЛУЧАЕТ ФАКТОР В, ВТОРОЙ ПЛОД ПОЛУЧАЕТ ФАКТОР А ВСЛЕДСТВИЕ АНАСТОМОЗА СОСУДОВ ПРОИСХОДИТ ОБМЕН КРОВЕОБРАЗУЮ-ЩИМИ КЛЕТКАМИ, НО ОБА ПЛОДА ИМЕЮТ ОДИНАКОВЫЙ ТИП КРОВИ БЛИЗНЕЦЫ ИМЕЮТ ИДЕНТИЧНЫЙ ТИП КРОВИ (АВ) ОДНОЯЙЦОВЫЕ БЛИЗНЕЦЫ  БЛИЗНЕЦЫ ИМЕЮТ СМЕСЬ КРОВИ ДВУХ ТИПОВ: ТИП А (БЕЗ В) И ТИП В (БЕЗ А) Рис. 43. Схема анастомоза сосудов у двуяйцевых близнецов (справа), вызывающего мозаичное» эритроцитов. У однояйцовых близнецов, несмотря на анастомоз сосков, тип крови всегда одинаковый (по И. Иоганеону и др., 1970) 216 антигены друг друга (явление толерантности), поэтому в течение всей жизни можно проводить (как и у однояйцовых близнецов) пересадку органов и тканей. Около 90 % телок из разнополых двоен в результате анастомоза сосудов становятся бесплодными — фримартинами, и их, естественно, приходится выбраковывать. Сейчас существует точка зрения, что антиген Н—Y направляет развитие недифференцированных гонад по мужскому (тестикулярному) типу. Бесплодие самок вызвано не передачей тестостерона от бычка-близнеца телочке, как предполагали раньше, а химерностью половых хромосом у самки (XX/XY). Развитие в химерных яичниках клеток XX по мужскому типу определяется антигеном Н—Y, который вырабатывается клетками XY. С помощью групп крови можно выявить до 98 % дизиготных пар. Химеризм эритроцитов встречается у человека (очень редко), овец и свиней. Межпородная и внутрипородная дифференциации. Группы крови, как и другие биохимические полиморфные системы, позволяют изучать историю эволюции домашних животных, происхождение и родство пород, генетическую структуру их и внутри-породную дифференциацию, проводить планирование и контроль селекционного процесса. Одна из самых жирномолочных пород мира — джерсейская имеет ряд В-аллелей, которые не встречаются у других западноевропейских пород скота, кроме гернсейской. У этой породы также высока частота антигена Z', тогда как у западноевропейских пород он редок, но зато встречается у зебу Африки, Азии и скота юга Восточной Европы. Подтверждено с помощью групп крови генеалогическое родство голландского и холмогорского скота. В. Н. Тихонов (1991) установил, что антиген Fa встречается почти у всех животных вьетнамской черной, польско-китайской и беркширской пород свиней (около 100 %), в меньшей степени у кемеровской, миргородской и северокавказской (54 и 38 %), низкая частота у украинской степной (3 %), тогда как у свиней крупной белой, эстонской белой и других пород этот антиген отсутствует или имеет очень низкую частоту. Эти исследования объяснили филогенез многих современных пород от древних свиней Юго-Восточной Азии и показали генетическое сцепление локусов систем групп крови F с локусом белой масти. Выявлена и внутрипородная дифференциация животных по группам крови в пределах линий и семейств. Ряд ученых указывают на возможность поддержания генетического сходства животных линий с родоначальником и выведения генетически маркированных линий животных с использованием групп крови. Построение генетических карт. Изучение сцепления локусов групп крови и биохимических полиморфных систем и частоты кроссинговера между ними дает возможность составить генетические карты хромосом. Карты хромосом позволяют следить за 217 ПЛЕЙОТРОПИЯ МАРКЕРНЫЙ ГЕН РЕЗИСТЕНТНОСТЬ ИЛИ восприимчивость к БОЛЕЗНИ ВНЕШНЯЯ СРЕДА (ПИТАНИЕ, КЛИМАТ, ИНФЕКЦИИ, ПРОДУКТИВНОСТЬ и тд) ГРУППЫ КРОВИ, БИОХИМИЧЕСКИЕ ПОЛИМОРФНЫЕ СИСТЕМЫ ХРОМОСОМЫ \ \ \ ГЕНЫ СЦЕПЛЕНИЕ ГЕН РЕЗИСТЕНТ-НОСТИ ИЛИ ВОСПРИИМЧИВОСТИ РЕЗИСТЕНТНОСТЬ ИЛИ ВОСПРИИМЧИВОСТЬ К БОЛЕЗНИ ВНЕШНЯЯ СРЕДА МАРКЕРНЫЙ ГЕН 11 f ГЕНЫ ГЕТЕРОЗИС наследственной передачей болезней, если они сцеплены с группами крови или другими полиморфными системами. У свиней J- и С-локусы групп крови сцеплены с генами главного локуса гистосовместимости свиней (SLA). Частота кроссинговера между J- и С-локусами равна 6 сМ, а между J-локусом и SLA — 9,8 сМ. В. Н. Тихонов и др. (1982) впервые картировали локус структурного гена аллотипа А=23 альфа-глобулина сыворотки крови в хромосоме № 16 свиньи. В. Н. Тихоновым обобщены данные по картированию большой группы локусов у свиней (рис. 44). Определено расстояние локусов от центромеры. По мнению других авторов, J-, С- и SLA-локусы находятся в 7-й хромосоме, локус группы крови Н, контролирующий ингибицию А- и О-факторов (S) и чувствительность к галотану (Hal), — в 6-й хромосоме (P. Thomsen, 1990). Связь групп крови с резистентностыо к болезням. Известно, что заболеваемость язвой двенадцатиперстной кишки у людей с группой крови О (I) в 1,3—1,5 раза выше, чем у лиц с другими группами. Среди лиц с А (П)-группой частота пораженное™ туберкулезом и раком желудка в 1,4 и 1,2 раза соответственно больше, чем у лиц с О-группой. К 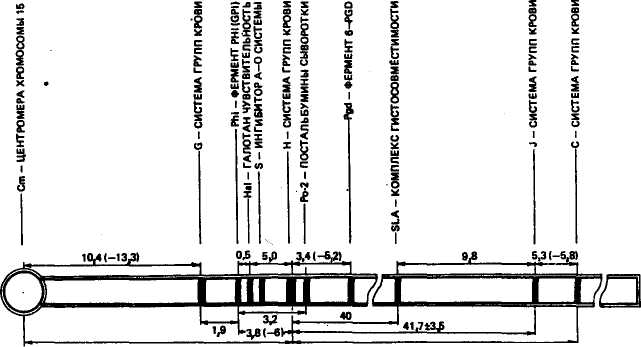 настоящему времени выполнено огромное количество работ по изучению корреляции групп крови и биохимических полиморфных систем с резистентностью к болезням, а также с различными признаками продуктивности. Поиск подобных связей основан на четырех (рис. 45) теоретических положениях: настоящему времени выполнено огромное количество работ по изучению корреляции групп крови и биохимических полиморфных систем с резистентностью к болезням, а также с различными признаками продуктивности. Поиск подобных связей основан на четырех (рис. 45) теоретических положениях: | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||