В. Н. Сайтаниди Рецензент членкорреспондент расхн в. Ф. Красота Петухов В. Л. и др. П31 Ветеринарная генетика В. Л. Петухов, А. И. Жигачев, Г. А. Назарова. 2е изд., перераб и доп. М. Колос, 1996. 384 с ил. Учебники
 Скачать 5.3 Mb. Скачать 5.3 Mb.
|

|
Глава 12 ГЕНЕТИКА ПОПУЛЯЦИЙ' Для улучшения племенных и продуктивных качеств животных необходимо знать генотипы не только отдельных индивидуумов, но и генетическую структуру всего стада или даже породы в целом. Важное значение для селекции имеют знания закономерностей наследственности и изменчивости в отсутствие и с учетом искусственного отбора и подбора животных, факторов, их определяющих. Исследования генетических процессов, протекающих в естественных условиях размножения животных, имеют большое значение для дальнейшего познания эволюции с целью управления этими процессами при разведении сельскохозяйственных животных. ПОПУЛЯЦИЯ И «ЧИСТАЯ ЛИНИЯ» По Н. В. Тимофееву-Ресовскому, популяция — это совокупность особей данного вида, в течение длительного времени (большого числа поколений) населяющая определенное пространство, состоящая из особей, могущих свободно скрещиваться друг с другом, и отделенная от таких же соседних совокупностей одной из форм изоляции (пространственной, сезонной, физиологической, генетической). Например, олени острова Колгуев изолированы от оленей, разводимых на материковой части Крайнего Севера, широкой полосой моря. В результате сформировалась особая популяция колгуевских оленей, отличающаяся от другой части этого вида генотипическими и фенотипическими признаками — они более крупные и обладают лучшей жизнеспособностью. В животноводстве под популяцией понимают группу животных одного вида, характеризующихся определенной численностью и ареалом распространения. Такая группа отличается от других популяций генетической структурой, экстерьерными, ин-терьерными и продуктивными качествами. Популяцией в животноводстве может быть отдельное стадо животных, порода или отродье. Обычно популяция — замкнутая группа. Ввоз в нее или вывоз из нее животных из других популяций ограничен, поэтому размножение в популяции осуществляется за счет подбора сам- 196 цов и самок, принадлежащих к данной популяции. В Ярославской области, например, разводится популяция крупного рогатого скота ярославской породы. Каждая популяция характеризуется определенным генофондом, т. е. совокупностью аллелей, входящих в ее состав. Наряду с популяцией в генетике существует понятие «чистая линия» — это потомство, полученное только от одного родителя и имеющее с ним полное сходство по генотипу. Чистые линии могут быть созданы в растениеводстве у самоопыляющихся растений. В отличие от популяций они характеризуются полной гомозиготностью. Вследствие полной гомозиготности отбор в чистой линии невозможен, так как все особи, входящие в нее, имеют идентичный набор генов. Высокогомозиготных линейных мышей, крыс и других лабораторных животных создают в целях проведения различных экспериментов, например для проверки на мутагенность тех или иных препаратов, оценки вакцин и т. д. Популяция состоит из животных разных генотипов. Эффективность отбора в ней зависит от степени генетической изменчивости — соотношения доминантных и рецессивных генов. СТРУКТУРА СВОБОДНО РАЗМНОЖАЮЩЕЙСЯ ПОПУЛЯЦИИ. ЗАКОН ХАРДИ - ВАЙНБЕРГА Харди и Вайнберг провели математический анализ распределения генов в больших популяциях, где нет отбора, мутаций и смешивания популяций. Они установили, что такая популяция находится в состоянии равновесия по соотношению генотипов, что определяется формулой р*АА + + 2pqAa + q^aa = 1, где р — частота доминантного гена А, q — частота его рецессивного аллеля а. В соответствии с этим был сформулирован закон, или правило, Харди — Вайнберга, согласно которому при отсутствии факторов, изменяющих частоты генов, популяции при любом соотношении аллелей от поколения к поколению сохраняют эти частоты аллелей постоянными. Несмотря на известные ограничения, по формуле Харди — Вайнберга можно рассчитать структуру популяции и определить частоты гетерозигот (например, по летальным или сублетальным генам, зная частоты гомозигот по рецессивным признакам и частоты особей с доминантным признаком), проанализировать сдвиги в генных частотах по конкретным признакам в результате отбора, мутаций и других факторов. Пользуясь формулой Харди — Вайнберга, мы рассчитали частоту гетерозиготных носителей некоторых форм рецессивно обусловленных аномалий в стаде крупного рогатого скота племзаво-да «Караваево» (табл. 28). 197 Г?НОВ 28. Частота гетерозиготных носителей Р«ньк «^ ■ костромской породе (проанализировано 9608 Частота гетерозиготных носителей в популяции (Аа), % Число случаев уродств (да) Ф  енотипическое проявление енотипическое проявление11,2 3,9 4,3 3,3 3,3 36 4 5 3 3 У корочение нижней челюсти Мопсовидность Водянка плода Атрезия ануса Слепота Популяция находится в равновесии только тогда, когда в ней не происходит отбора. При выбраковке же отдельных животных в такой популяции изменяется соотношение гамет, что влияет на генетическую структуру следующего поколения. Однако К. Пирсон показал, что, как только возникает состояние панмиксии (свободное скрещивание), соотношение генотипов и фенотипов в популяфш в следующем поколении возвращается к тому, которое соответствует формуле Харди — Вайнберга, но уже при другом их соотношении. Скрещивание, восстанавливающее соотношение генотипов в популяции, в соответствии с формулой Харди — Вайнберга получило название стабилизирующего. Из этого следует вывод: при использовании в популяции случайных, неотобранных производителей или маток наблюдается стабилизация признаков продуктивности на одном уровне, и повышение продуктивности животных в такой ситуации невозможно. Точно так же при отсутствии браковки гетерозиготных носителей рецессивных аномалий частота проявления аномальных животных в популяции остается неизменной. ОСНОВНЫЕ ФАКТОРЫ ГЕНЕТИЧЕСКОЙ ЭВОЛЮЦИИ В ПОПУЛЯЦИЯХ В популяциях сельскохозяйственных животных постоянно изменяются частоты генов, что можно наблюдать при анализе смежных поколений. Такие изменения составляют суть генетической эволюции. Основные факторы эволюции: мутации, естественный и искусственный отбор, миграции, дрейф генов. Одна из основных причин генетической изменчивости в популяции — мутации. Спонтанные мутации каждого гена происходят с низкой частотой, однако общая частота мутаций всех генов, которые содержат популяции, очень велика. Мутации, возникающие в половых клетках родительского поколения, приводят к изменению генетической структуры у потомства. В популяции постоянной численности в отсутствие отбора большинство возникших мутаций быстро утрачивается, однако некоторые из них могут сохраниться в ряде поколений. Исчезновению мутантных 198 генов из популяции противостоит действие мутационного процесса, в результате которого образуются повторные мутации. Генетическая структура популяций формируется и изменяется под действием естественного и искусственного отбора. Действие естественного отбора состоит в том, что преимущественное размножение имеют особи с высокой жизнепособностью, скороспелостью, плодовитостью и т. д., т. е. более приспособленные к условиям окружающей среды. При искусственном отборе определяющее значение имеют признаки продуктивности. В. И. Власов отмечает, что естественный отбор идет на всех этапах онтогенеза популяции — от образования гамет до взрослого организма. При этом он существенно влияет на темпы искусственного отбора вследствие противоположного действия при селекции на высокий уровень развития продуктивных признаков, несвойственный видовым биологическим границам. Исходя из этого, при отборе животных необходимо учитывать не только продуктивные признаки, но и признаки приспособленности к условиям окружающей среды. По С. М. Гершензону, критерием интенсивности естественного отбора служит разность приспособленности сравниваемых групп, называемая коэффициентом отбора и выражаемая в долях единицы. Например, если вероятность оставления потомства особями с генотипом аа на 10 % меньше, чем особями с генотипом АЛ или Аа, то приспособленность этих групп для особей АА и Аа равна 1, для особей аа — 0,9. С точки зрения ветеринарной генетики имеет значение эффективность отбора против вредных мутаций, прежде всего рецессивного типа. Анализ показывает, что высокие частоты рецессивного мутантного гена путем отбора могут быть быстро снижены до низких значений. Чтобы снизить частоту летального гена, например с 0,3 до 0,2, достаточно двух поколений (табл. 29). 29. Снижение частоты рецессивного аллеля а при полной элиминации гомозигот aa (no Визнеру и Виллеру, 1979) Частота фенотипов Уменьшение q на одно поколение Ад Отношение 2РЯ-<Г Частота рецессивного аллеля 2pq Н еобходимое число поколений для
Из данных таблицы 29 видно, что частота гомозигот (аа) по мутантному гену зависит от частоты гетерозиготных животных в популяции. Выявление этих гетерозигот и элиминация их соот- 199 Степень родства родителей [ Уровень возрастания гомозиготности у потомства, % ветственно будут снижать частоту генетических аномалий, обусловленных мутантным геном, что особенно важно при высокой 30. Уровень возрастания roi при разных степенях инбридинга fteSiSS' структура популяции может изменяться в силу случайных генетико-автоматических процессов (по Н. 11. дуои-нину) или дрейфа генов (по С. Райту). Наблюдения показывают, что наиболее интенсивно дрейф генов протекает в малых популяциях Например, известны случаи высокой концентрации редких мутаций в малочисленных изолированных популяциях крупного рогатого скота и других видов животных, связанные, очевидно, с генетико-автоматическими процессами.Распространение мутаций в разных популяциях животных может произойти в результате миграций. На практике это означает завоз животных, особенно производителей или их спермы, из других зон страны или стран. Известно много случаев, когда импортные производители оказывались носителями мутации и распространяли генетические аномалии при использовании в воспроизйвдстве местных популяций. Так, экспорт голигганов из США в ФРГ по данным Рика, способствовал распространению пупочных грыж у немецкого черно-пестрого скота. В нашей стране наблюдалась ситуация, когда из одного племзавода ремонтных быков с транслокацией 1/29 хромосомы продавали без ограничений в разные области. Это могло бы привести к распространению нежелательной мутации в других популяциях. По рекомендации ученых были приняты меры по элиминации носителей транслокаций. ВЛИЯНИЕ ИНБРИДИНГА НА ВЫЩЕПЛЕНИЕ РЕЦЕССИВНЫХ ЛЕТАЛЬНЫХ И ПОЛУЛЕТАЛЬНЫХ ГЕНОВ Спаривание животных, находящихся в родственных отношениях называют инбридингом. Родственное спаривание, или инбридинг, -метод подбора, используемый в племенном животноводстве для закрепления ценных наследственных признаков того или иного животного в последующих поколениях. У родственных между собой животных наблюдается сходство по определенным парам аллелей, которые они получили от общего предка. Эю сходство тем больше, чем ближе степень Р°ДкХщое животное в генотипе имеет аллельные гены как в гомозиготном, так и в гетерозиготном состоянии. Ь гетерозиготе обычно находятся вредные мутантные рецессивные гены При инбридинге возрастает вероятность слияния тождественных гамет, несущих мутантные гены в гетерозиготном состоянии, и перехода их в гомозиготное COCT°^^pj™ вероятность пропорциональна степени родства спариваемых животных (табл. 30). 200 25,0 12,5 6,25 3,125 II—I II—II ш-ш ГУ—IV Повышение гомозиготности у потомства при разных степенях родственных отношений родителей определяют по несколько измененной формуле С. Райта: где Fx— коэффициент инбридинга потомка; я — рад родословной с материнской; /И — ряд родословной с отцовской стороны, где находится общий предок; fa — коэффициент инбридинга общего предка. Таким образом, в результате применения инбридинга происходит изменение генных частот, возрастает вероятность выщеп-ления рецессивных гомозигот, что является причиной инбредной депрессии, выражающейся в снижении жизнеспособности, плодовитости животных, рождении аномальных особей. Таблица 31 иллюстрирует пример перехода при инбридинге мутантных рецессивных генов в гомозиготное состояние и появление при этом различных форм уродств в потомстве быка костромской породы. 31. Формы уродств в потомстве быка Бурхана 6083 Число уродливых телят Форма уродства В  том числе инбредных на быков том числе инбредных на быковСалата Л  адка Бурхана* адка Бурхана*
* Инбридинг, как правило, был комплексным — одновременно на двух (1-я цифра) или на трех (2-я цифра) предков. Ниже показано, что инбредная депрессия может проявляться в форме снижения показателей продуктивности животных. Инбредная депрессия на каждые 10 % повышения коэффициента инбридинга следующая (по Ф. Хатту): 201 Изменения при инбридинге * -0,20 —0,44 -1,55 —4,4 +6,0 -9,0 Показателя продужпиности Свиньи: число поросят в помете при рождении число поросят в помете в возрасте 154 дней средняя масса одного поросенка в 154 дня, кг Куры: выводимость, % от числа оплодотворенных яиц возраст начала яйцекладки, дней число яиц, снесенных в среднем одной курицей к возрасту 500 дней Имбредная депрессия по показателям, характеризующим продуктивность и жизнеспособность животных, не является фатальным спутником родственного спаривания. Имеется множество примеров того, когда при инбридинге разных степеней, в том числе и близких, отрицательных последствий не наблюдали. Н. П. Дубинин в этой связи отмечает, что «линия ухудшается, пока в пей идут процессы последовательного накопления вредных рецессивных генов, переходящих в гомозиготное состояние. Когда же наступает более или менее выраженное завершение этого процесса, линии по своим свойствам становятся относительно константными и могут в таком устойчивом состоянии сохраняться длительно. Изменить генотип таких линий могут лишь новые накапливающиеся в них мутации». Однако, подчеркивает академик, «многие линии при инбридинге, конечно, гибнут, ибо в них в гомозиготное состояние переходят летальные и полулетальные гены». Поэтому инбридинг применяется как метод индивидуальной селекции для перевода в гомозиготное состояние ценных генов выдающихся животных. ГЕНЕТИЧЕСКИЙ ГРУЗ В ПОПУЛЯЦИЯХ ЖИВОТНЫХ В ходе длительной эволюции животных наряду с полезными мутациями, подхватываемыми отбором, в популяциях или породах накопился определенный спектр генных и хромосомных мутаций. Каждое поколение популяции наследует этот груз мутаций, и в каждом из них возникают новые мутации, часть которых передается последующим поколениям. Очевидно, что большая часть вредных мутаций отметается естественным отбором или элиминируется в процессе селекции. Это прежде всего доминантные генные мутации, фенотипически проявляющиеся в гетерозиготном состоянии, и количественные изменения наборов хромосом. Рецессивно действующие генные мутации в гетерозиготном состоянии и структурные перестройки хромосом, заметно не влияющие на жизнеспособность их носителей, могут проходить сквозь сито селекции. Они формируют генетический груз популяции. Таким образом, под генетическим 202 грузом популяции понимают совокупность вредных генных и хромосомных мутаций. Различают мутационный и сегрегационный генетический груз. Первый формируется вследствие новых мутаций, второй — в результате расщепления и перекомбинирования аллелей при скрещивании гетерозиготных носителей «старых» мутаций. Частота летальных, полулетальных и субвитальных мутантных генов, передающихся из поколения в поколение в форме мутационного генетического груза, из-за трудности идентификации носителей не поддается точному учету. Мортон и Кроу предложили форму расчета уровня генетического груза в количестве летальных эквивалентов. Один летальный эквивалент соответствует одному летальному гену, обусловливающему смертность с 10%-ной вероятностью, двум летальным генам при 50%-ной вероятности смерти и т. д. Величина генетического груза по формуле Мортона log eS = A + BF, где S— часть потомства, оставшаяся в живых; Л — смертность, измеряемая летальным эквивалентом в популяции при условии случайных спариваний (F= 0), плюс смертность, обусловленная внешними факторами; В — ожидаемое увеличение смертности, когда популяция становится полностью гомозиготной (F= 1); F — коэффициент инбридинга. Уровень генетического груза можно определять на основании фенотипического проявления мутаций (уродства, врожденные аномалии обмена и т. д.)> анализа типа их наследования, частоты в популяции. Н. П. Дубинин предлагает определять генетический груз популяции путем сравнения частот мертворожденных в родственных и неродственных подборах родительских пар. При этом следует иметь в виду, что при высокой частоте гетерозигот по рецессивным летальным и полулетальным мутантным генам рождение животных с аномалиями необязательно должно быть связано с инбридингом близких и умеренных степеней. Общий предок (источник мутации) может находиться и в отдаленных рядах родословной. К примеру, бык Трувор 2918 — гетерозиготный носитель мутантного рецессивного гена, находился в V, VI, VII рядах предков в совхозе «Красная Балтика», но при использовании его праправнука Автомата 1597 на родственных ему коровах наблюдались массовые случаи рождения бесшерстных телят (рис. 41). Другой прапраправнук Трувора бык Док 4471 также оказался гетерозиготным носителем гена бесшерстности. В совхозе «Новое время» при умеренно родственных спариваниях и отдаленном инбридинге в потомстве Дока 4471 зарегистрировано примерно 5 % телят с этой генетической аномалией. Эти данные в определенной мере характеризуют уровни гене- 203 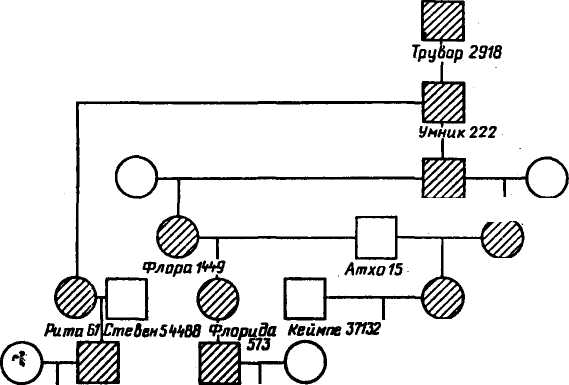  о Лукабый 1BJL Динка1Б15 Динка 1В32 Динка 256В Финка 1262 Студент ПЗ Рая 137В \ Глазурь 1172 дак¥П1 Рис. 41. Схема родственных связей родителей телят черно-пестрой породы с синдромом бесшерстности, укорочения верхней челюсти, «волчьей пасти» тического груза по отдельным мутантным генам в конкретных популяциях крупного рогатого скота. Хромосомные мутации являются составной частью генетического груза. Учет их ведется прямым цитологическим методом. По результатам многочисленных исследований основной компонентой груза аберраций хромосом у крупного рогатого скота являются робертсоновские транслокации, а у свиней — реци-прокные. Наиболее распространенной мутацией у крупного рогатого скота оказалась транслокация 1/29 хромосомы. Размах изменчивости частоты этой аберрации, по нашим данным, в популяциях палево-пестрого скота составлял от 5 до 26 %. Таким образом, концепция генетического груза в свете современных достижений цитогенетики должна быть расширена. Сейчас, когда известен широкий спектр аберраций хромосом и уста- 204 новлено строгое наследование отдельных из них (транслокации и инверсии), представляется целесообразным учитывать их наряду с вредными мутациями генов как составляющую часть генетического груза. ГЕНЕТИЧЕСКАЯ АДАПТАЦИЯ И ГЕНЕТИЧЕСКИЙ Г0МЕ0СТАЗ ПОПУЛЯЦИЙ Существование в популяциях наследственной изменчивости, прежде всего мутаций в гетерозиготном состоянии, позволяет им быстро приспосабливаться к новым условиям среды за счет изменения генетической структуры. Мутационный процесс ведет также к образованию в популяциях генетического полиморфизма — разнообразия частот аллелей, гомозигот по доминантным, гетерозигот или гомозигот по рецессивным генам. Полиморфизм является механизмом, поддерживающим существование популяций. Если, например, гетерозиготность обеспечивает лучшую приспособляемость к изменившимся условиям среды, то идет отбор в пользу гетерозигот, что приводит к сбалансированному полиморфизму — воспроизведению в популяции из поколения в поколение определенного соотношения различных генотипов и фенотипов. Процессы, обеспечивающие способность популяции сохранять свою генетическую структуру, называют генетическим гомеостазом. ОСОБЕННОСТИ НАСЛЕДОВАНИЯ КОЛИЧЕСТВЕННЫХ ПРИЗНАКОВ В генетике выделяют два класса признаков — качественные и количественные. Они различаются по характеру изменчивости и особенности наследования. Качественные признаки характеризуются прерывистой, а количественные — непрерывной изменчивостью. Первые из них дают четкие границы при расщеплении на доминантные или рецессивные признаки. Это связано с тем, что каждый из них обычно контролируется одним аллельным геном. Количественные признаки не дают четких границ расщепления при разных вариантах скрещивания, хотя отличаются от качественных более высокой степенью изменчивости. Особенностью количественных признаков является сложный характер наследования. Каждый из них детерминируется не одним, а множеством локусов в хромосомах. Такой тип наследования, когда один признак обусловливается многими генами, носит название полигенного. Уровень развития количественного признака зависит от соотношения доминантных и рецессивных генов, других генетических факторов и степени модифицирующего действия факторов внешней среды. Изменчивость по количественному при- 205 знаку в популяции складывается из генетической и паратипичес-кой (внешнесредовой) изменчивости. Понятие о наследуемости признаков и коэффициенте наследуемости. Разные количественные признаки имеют неодинаковую степень генетической изменчивости, и условия внешней среды оказывают различное воздействие на уровень фенотипического проявления того или иного признака. При отборе животных важнейшее значение имеет знание того, в какой степени будет совпадение уровней развития количественного признака у родителей и потомства или в какой степени потомки унаследуют количественные хозяйственно полезные признаки или патологические признаки родителей. Для оценки эффективности отбора по количественным признакам в популяционной генетике введено понятие «наследуемость». Под наследуемостью признака понимают степень его генетической детерминации в фенотипе. Другими словами, определяют, в какой степени уровень развития признака зависит от генотипа* родителей и в какой степени — от условий внешней среды. Коэффициент наследуемости (А2) указывает на долю генетической изменчивости признаков в общей изменчивости. Наиболее распространенными методами вычисления А2 являются следующие: 1) А2 = 2гд/м, где г— корреляция по количественному признаку между прямыми родственниками, например мать — дочь; 2) А2 = 2R д/м, где R — коэффициент регрессии между фенотипами прямых родственников. В последнее время получило распространение мнение 6 том, что отдельные качественные признаки имеют сложный полигенный характер наследования. К этим признакам относят и устойчивость к болезням. Для качественных признаков используют метод вычисления величины А2 через коэффициент регрессии. Д. В. Карликов разработал полигенную модель наследования устойчивости к лейкозу и рассчитал коэффициент наследуемости по этому признаку по формуле А2= Ьг, где г — коэффициент родства больных животных с их родственниками; Ъ — регрессия, определяемая по отношению Ь = (R— G):(A— G). Здесь R — средняя подверженность родственников больных животных; G — средняя подверженность популяции к лейкозу, А — средняя подверженность больных животных в популяции. Наследуемость хозяйственно полезных признаков у животных. К хозяйственно полезным признакам относят молочность," со- 206 держание жира и белка в молоке коров, настриг шерсти у овец, яйценоскость у кур, прирост живой массы, плодовитость и др. Повышение уровня развития хозяйственно полезных признаков достигается постоянным отбором лучших индивидуумов для воспроизводства. Эффективность отбора по этим признакам зависит от степени их наследуемости, взаимосвязи между ними, разности между средним значением признака отобранной группы и средним по стаду (селекционный дифференциал) и интервала между поколениями. Величины коэффициентов наследуемости зависят от природы признака. Так, среднее значение А2 по удою равно 0,25, жирномолочности — 0,38, живой массе у овец — 0,35, выходу чистой шерсти — 0,55, плодовитости у крупного рогатого скота — 0,08 и т. д. ГЕНЕТИЧЕСКИЕ ОСНОВЫ ГЕТЕРОЗИСА Селекционной практикой и специальными исследованиями накоплены данные, свидетельствующие о том, что в некоторых случаях уровень развития одного или нескольких признаков у потомства превосходит степень выраженности этих признаков у лучшего из родителей. Такое явление, названное гетерозисом, не вписывается в обычные рамки наследования признаков. Для его объяснения предложены разные гипотезы: 1) гетерозиготного состояния по многим генам; 2) взаимодействия доминантных благоприятных генов; 3) сверхдоминирования, когда гетерозиготы превосходят гомозиготы. Н. В. Турбин предложил теорию генетического баланса, в основе которой лежит сложный характер причинно-следственных связей между наследственными факторами и признаками. Н. Г. Дмитриев и И. JI. Гальперн отмечают, что главную причину возникновения гетерозиса надо искать в особенностях эволюции вида, породы, линии. При этом следует иметь в виду, что все в природе направлено на сохранение жизни. Проявление гетерозиса зависит от генетической природы признака. Так, при Межпородном или межлинейном скрещивании гетерозис в большей степени проявляется в отношении признаков, имеющих Низкую степень наследуемости. Что касается признаков, имеющих среднюю или высокую наследуемость, то гетерозис по ним Чаще всего проявляется слабо, и гибриды занимают обычно промежуточное положение. Для расчета гетерозиса рекомендуется использовать формулу К. Б. Свечина: „Г_ где ЕП — показатель признака у гибрида Fj; Ep— показатель этого же признака У лучшей родительской формы. 207 Н. Г. Дмитриев и И. Л. Гальперн предлагают еще одну формулу вычисления- гетерозисного эффекта: F ИГ= 1 где F\ — средний показатель продуктивности гибридов; Afp — лучший показатель родительской формы. При наличии истинного гетерозиса величина индекса или больше 100 %, или имеет знак «плюс». Если же величина гетерозиса меньше 100 % или имеет знак «минус», то правильнее говорить о лучшей или худшей комбинационной способности линий. Скрещивание последних по определенной схеме обеспечивает в гибридном потомстве лучшее развитие одного признака от отца, а другого от матери, хотя этот признак у гибрида по своему развитию не превосходит лучшую родительскую форму. вопросы. 1. Что такое популяция и «чистая линия»? 2. В чем сущность и практическое значение закона Харди — Вайнберга? 3. Какие факторы определяют генетическую эволюцию в популяциях? 4. Какова роль инбридинга в появлении уродств у животных и других генетических аномалий? 5. В чем сущность концепции генетического груза? Как определить уровень генетического груза в популяции? 6. Каковы особенности наследования количественных признаков? 1 |