|
В. Н. Сайтаниди Рецензент членкорреспондент расхн в. Ф. Красота Петухов В. Л. и др. П31 Ветеринарная генетика В. Л. Петухов, А. И. Жигачев, Г. А. Назарова. 2е изд., перераб и доп. М. Колос, 1996. 384 с ил. Учебники
42^*3,0
14,31-18,51
Рис. 44. Генетическая карта 15-й хромосомы свиньи (по Тихонову, 1984). Все расстояния от центромеры до локусов и между локусами указаны в сантиморгаиах (сМ). (Использованы данные Андерсена, 1970; Расмусена н др., 1982; Иоргенсева н др., 1981; Хрубана и др., 1976; Джунеа и др., 1982)
218
РЕЗИСТЕНТНОСТЬ ИЛИ
ВОСПРИИМЧИВОСТЬ К
БОЛЕЗНИ (
МАРКЕРНЫЙ ГЕН
ВНЕШНЯЯ СРЕДА
( ГЕТЕРОЗИГОТА) АА < At >м ГЕТЕРОЗИГОТА) АА < At >м
t t t ГЕНЫ
АНТИГЕНЫ
А,
'ft
ИММУНОЛОГИЧЕСКАЯ НЕСОВМЕСТИМОСТЬ "МАТЬ-ПЛОД"
МАТЬ АНТИГЕНЫ ПЛОД НОВОРОЖДЕННЫЙ
Т    ||ТИП АНТИТЕЛА (АНТИ-А,) ||ТИП АНТИТЕЛА (АНТИ-А,)
Р ис. 45. Генетические причины взаимосвязи групп крови, биохимических полиморфных систем с резистентностью, восприимчивостью к болезням и продуктивностью ис. 45. Генетические причины взаимосвязи групп крови, биохимических полиморфных систем с резистентностью, восприимчивостью к болезням и продуктивностью
плейотропном действии генов, т. е. когда гены,
обусловливающие группы крови или биохимические полиморф
ные системы (маркерные гены), прямо или косвенно влияют на
резистентность к болезням и продуктивность;
сцеплении между локусами групп крови или биохими
ческих полиморфных систем и локусами, влияющими на резис
тентность или продуктивность;
гетерозисе, когда гетерозиготность по группам крови
или биохимическим полиморфным системам повышает резис
тентность к болезням или продуктивность;
иммунологической несовместимости ма
тери и плода, при которой вследствие разных генотипов у матери
и плода по группам крови возникают, например, гемолитическая
болезнь у жеребят, поросят, эритробластоз у человека.
219
Н-группа крови используется для определения чувствительности свиней к синдрому стресса (PSS), который характеризуется внезапной смертью животных, вызванной транспортировкой, высокой температурой и другими стрессорами. К PSS чувствительны гомозиготные НаНа особи. Локусы Н-системы группы крови и PHI (фосфогексоизомеразы) связаны с чувствительностью к синдрому злокачественной гипертермии (MHS), который вызывается лекарственными веществами, галотаном.
Аллель В21 группы крови у птиц коррелирует с повышенной резистентностью к болезни Марека. Цыплята генотипа В2/В2 более резистентны к вирусу саркомы Рауса, чем особи с генотипом В5/В5.
Гемолитическая болезнь новорожденных. В 1940 г. Левин с сотрудниками открыли гемолитическую болезнь новорожденных у человека, обусловленную несовместимостью генотипов матери и плода. В браках резус-положительных (Rh+) мужчин с резус-отрицательными (Rh) женщинами могут рождаться резус-поло-жителв&ые дети. На 2—3-м месяце беременности кровь резус-положительного плода, поступая в организм матери, вызывает образование у нее антител против резус-антигена. Антитела, проникая через плаценту в организм плода, вызывают эритроб-ластоз (разрушение эритроцитов).
ПЛАЦЕНТА
ПЛОД
ПЛОДОВЫЕ ОБОЛОЧКИ С КРОВЕНОСНЫМИ СОСУДАМИ
АНТИТЕЛА
ЭРИТРОЦИТЫ
В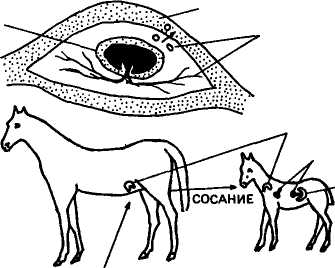 о многом сходное заболевание встречается у поросят, жеребят и телят. Но в отличие от человека плацента указанных видов непроницаема для антител и они накапливаются в молозиве (рис. 46). Только после сосания матери в первые 24—48 ч у о многом сходное заболевание встречается у поросят, жеребят и телят. Но в отличие от человека плацента указанных видов непроницаема для антител и они накапливаются в молозиве (рис. 46). Только после сосания матери в первые 24—48 ч у
РЕАКЦИЯ АНТИГЕН-ЭРИТРОЦИТ - АНТИТЕЛО
молозиво с
АНТИТЕЛАМИ
Рис. 46. Развитие гемолитической болезни у жеребят:
А — эритроциты плода попадают через плаценту и кровоток матери; Б — образовавшиеся в крови антитела поступают с молозивом в организм жеребенка, вызывая разрушение эритроцитов
220
новорожденного наблюдаются патологические изменения в виде желтушности склеры глаз, слабости, учащенного дыхания, снижения числа эритроцитов. Молодняк в таких случаях погибает в течение нескольких дней.
У лошадей изогемолиз новорожденных наиболее часто возникает, когда жеребята имеют Ai- и Q-антигены соответствующих систем групп крови, наследуемых от отца и отсутствующих у матерей. Иногда иммунологический конфликт наступает при наследовании потомков от отца антигенов R и S. Своевременное незадолго до выжеребки выявление антител у матерей и поение жеребенка первые два дня жизни молозивом другой кобылы позволяют избежать заболевания. В это время молозиво матери сдаивают.
Частота изогемолиза новорожденных у жеребят английской чистокровной породы составляет около 1 %. Полагают, что эта болезнь в основном встречается у лошадей арабской породы и других, от нее происходящих.
Естественный изогемолиз новорожденных у крупного рогатого скота встречается редко, поэтому до 1970 г. не было зарегистрировано ни одного случая заболевания. В настоящее время имеется много данных о том, что в стадах, вакцинированных против анаплазмоза, частота изогемолиза (N1) достигает 3—20 %. По данным Керр (1973), в одном стаде от 24 коров, за год до отела вакцинированных против анаплазмоза, было 66,6 % пораженных N1 телят, из которых 18 % погибло. Полагают, что в большинстве случаев изогемолиз новорожденных у крупного рогатого скота — следствие вакцинации против анаплазмоза.
У свиней, как и у лошадей, основная причина N1 — несовместимость по группам крови матери и плода.
Связь групп крови с продуктивностью. Селекционеры давно мечтают найти маркеры для прогнозирования продуктивности в раннем возрасте. Удобно было бы использовать в качестве генетических маркеров группы крови и биохимические полиморфные системы. Много сил потрачено на изучение этой проблемы, но и сегодня она далеко не решена.
У шведского черно-пестрого и красно-пестрого скота выявлена положительная корреляция аллеля BO1Y2D' системы В с содержанием жира в молоке. Л. К. Эрнст и др. (1973) показали, что аллель 12 В-системы связан с жирномолочностью коров ряда линий черно-пестрой и ярославской пород. По данным В. Ф. Красоты, коровы костромской породы с некоторыми аллелями (О, Р) В-системы отличались более высокой молочностью. Аллели В1 и В3 у кур коррелируют с высокой яйценоскостью.
Повышение продуктивности может быть связано и с гетерози-готностью по группам крови. Так, увеличение гетерозиготности по В-локусу у кур привело к повышению вылупляемости цыплят, интенсивности роста и яйценоскости.
Одна из гипотез, объясняющих гетерозис (превосходство гиб-
221
Продолжение
Число аллелей
Символ локуса
ридов над родительскими формами по степени развития того или иного признака), — гипотеза сверхдоминантнрсти. Она основывается на утверждении, что в гетерозиготе 'гены более полно проявляются, чем в гомозиготе. В. Н. Тихонов установил, что гетерозиготность по некоторым антигенам групп крови ведет к гетерозису. При спаривании гомозиготных особей типа Gbb x х Gbb в среднем от свиноматки получено 10,67 поросенка, при спаривании гетерозиготных животных типа Gab x Gab — 11,47, а при спаривании Gaa x Gbb — 12,34 поросенка (гетерозис по плодовитости). В последнем случае масса гетерозиготных поросят в 2-месячном возрасте выше на 11 %.
БИОХИМИЧЕСКИЙ ПОЛИМОРФИЗМ
В течение эволюции в результате мутаций изменяются гены, поэтому в популяции они встречаются не в одной, а в двух и более формах (множественные аллели). Полиморфизм — одновременное присутствие двух или более генетических форм одного вида в таком численном отношении, что их нельзя отнести к повторным мутациям. Поэтому термин «генетический (биохимический) полиморфизм» применяется в тех случаях, когда локус хромосомы в популяции имеет два и более аллелей с частотой больше 0,01. Ген, представленный более чем одним аллелем, называют полиморфным геном. Доля полиморфных локусов точно неизвестна, но полагают, что в популяциях многих видов она достигает 25—50 %. Так, у человека из 50 тыс. или более структурных локусов по крайней мере 30 % могут быть полиморфными.
Основными методами изучения полиморфизма белков и ферментов являются электрофорез в крахмальном или акриламид-ном геле и иммуноэлектрофорез. Белки (в том числе ферменты) находятся в растворе в виде частиц, несущих определенный электрический заряд, которые под действием электрического тока перемещаются к катоду или аноду.
Сейчас у сельскохозяйственных животных изучено более 150 полиморфных локусов белков (в том числе ферментов) крови, молока, тканей (табл. 34), расположенных в аутосомах. Установлено сцепление трех локусов казеина молока ocSi-Cn, P-Cn и к-Cn (каппа-казеин).
34. Некоторые биохимические полиморфные системы
Символ локуса
Ч исло аллелей исло аллелей
Система
Свиньи
Овцы
Лошади
Куры
Крупный рогатый скот
Гемоглобин
Альбумин
Трансферрин
НЬ Alb Tf
5 7 13
2 3 10
2 3 4
222
10 9 12
Система
Свиньи
Овцы
Лошади
Куры
Крупный рогатый скот
Цсрулоплазмин
|
Ср
|
3
|
Эстераза
|
Es
|
2
|
р-Лактоглобулин
|
P-Lg
|
4
|
as 1-Казеин
|
asj-Cn
|
4
|
р- Казеин
|
P-Cn
|
6
|
к-Казеин
|
к-Cn
|
2
|
у-Казеин
|
y-Cn
|
4
|
Аллели гемоглобинового локуса обозначаются так: HtA HbB и т. д., а генотип — Hb^HtA Hb°HbB и т. д. В связи с кододоми-нантным наследованием большинства биохимических систем фенотип животного соответствует его генотипу, поэтому фенотип можно записать НЬАА или НЬА, НЬВВ или НЬВ.
Замещение аминокислот в белке может вызвать функциональные различия полиморфных форм. Например, у человека кроме нормального гемоглобина НЬ* известно более 50 патологических вариантов S, С, G и т. д., которые вызывают различные гемоглобинопатии (серповидно-клеточная анемия S, талассемия С). Одним из первых был открыт гемоглобин серповидных эритроцитов, который от нормального отличается заменой в шестом положении глутаминовой аминокислоты на валин. В районах распространения тропической малярии лица, гомозиготные по HbsHbs, погибают в раннем возрасте от серповидно-клеточной анемии. Гетерозиготы МУ^НЬ8 устойчивы к малярии, а люди с нормальным генотипом НЬАНЬА предрасположены к заболеванию.
Это неоспоримый пример сбалансированного полиморфизма, когда приспособленность гетерозигот выше, чем гомозигот, а оба аллеля сохраняются в популяции с промежуточной частотой. Это доказывает существование однолокусного гетерозиса по устойчивости к болезни. В. П. Эфроимсон (1968) выдвинул гипотезу о том, что иммунитет к малярии имеет адаптивное значение и обусловлен изменением молекулы гемоглобина НЬ, что препятствует его использованию малярийным плазмодием.
Гемоглобин выполняет важную для организма функцию переноса кислорода из органов дыхания к тканям и переноса углекислого газа от тканей в органы дыхания. У крупного рогатого скота открыто 10 типов гемоглобина, но у скота швицкой, костромской, джерсейской и других пород в основном встречаются аллели ШЛ и НЬВ. У животных черно-пестрой, айрширской, герефордской и других пород имеется только один тип А.
Хорошо изучен полиморфизм трансферрина (Tf), который переводит железо плазмы в диионизированную форму и
223
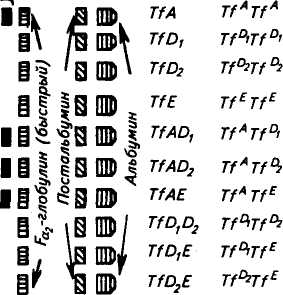
переносит его в костный мозг, где оно используется вновь для кроветворения. Трансферрин также подавляет размножение вирусов в организме. У человека недостаточность трансферрина может быть следствием некоторых перенесенных заболеваний, в частности наследственного гемохроматоза. Количество Tf снижается при циррозе печени, инфекционных болезнях. На рисунке 47 представлена схема расшифровки электрофореграммы типов трансферрина. Известно 12 аллелей Tf, но среди европейских пород наиболее часто встречаются аллели A, Di, D2 и Е.
Белок церулоплазмин (Ср) играет центральную роль в обмене меди в организме, являясь основным переносчиком ее в ткани. Нарушение функции церулоплазмина или снижение его содержания в плазме крови ведет, например у человека, к возникновению генетического заболевания нервной системы с некротическими изменениями в печени.
Все больше появляется работ по иммуногенетическому анализу белковых систем. Генетически детерминируемые анти-генные*^варианты сывороточных белков, по которым различают особей одного вида, называют аллотипами. О. К. Баранов (1981)" у американской норки выявил 8 аллотипов липопротеина (Lpm), обозначенных цифрами от 1 до 8. Липопротеины транспортируют липиды. Предполагают, что аллотипы Lpm-системы кодируются комплексом тесно сцепленных гомологичных генов. Аллотипы в основном наследуются аллогруппами,
Миграция
Старт
(+J Фенотип Генотип
Трансферрин
i
а! I
i i i
:1
!0f
1 i i i i i i i i i i ii i i i
£ D2 D, С
В А
Рис. 47. Расшифровка электрофореграммы различных типов сывороточных трансферринов крупного рогатого скота
224
I
например Lpm6>8, Lpm4>6> 8, Lpm3>4>6>8 и т. д. Аллогруппа — совокупность аллотипов, наследуемых как одна группа. Совокупность сцепленных генов одной хромосомы, контролирующих аллогруппу, называют гаплотипом.
У свиней идентифицированные аллотипы липопротеина детерминируются генами пяти локусов, временно обозначенных р, г, s, t, u. Закрытая система Lpb включает 8 аллелей, Lpr и Lpu — по два аллеля, а открытые системы Lps и Lpt — один аллель. Все аллотипы определяются аутосомными кодоминантными генами. Локусы и, р, t тесно сцеплены, а г и s локализованы в разных хромосомах. Имеются данные о связи некоторых типов Lpb с артериосклерозом у свиней.
ЗНАЧЕНИЕ БИОХИМИЧЕСКОГО ПОЛИМОРФИЗМА
Биохимические полиморфные системы белков используются для следующих целей:
изучения причин и динамики генотипической изменчивос
ти, составляющей основу эволюционной генетики;
уточнения происхождения отдельных животных;
описания межпородной и внутрипородной дифференциа
ции, изучения филогенеза и аллелофонда пород, линий и се
мейств, а также генетических процессов, происходящих в попу
ляциях животных, и изменения их генетической структуры в
процессе селекции;
определения моно- и дизиготных двоен;
построения генетических карт хромосом;
подбора гетерозисной сочетаемости;
выявления связи с резистентностью к заболеваниям, про
дуктивностью;
использования биохимических систем в качестве генетичес
ких маркеров в селекции животных.
Изучение 9 полиморфных систем белков у 10 главных групп скота позволило подтвердить вывод о том, что зебувидный скот Индии значительно отличается от европейских пород и принадлежит к другому виду (Bos indicus). Санга (тип африканского горбатого скота) занимает промежуточное положение между индийским зебу и европейскими породами, но в то же время имеет свои уникальные признаки. Часть из них — следствие обмена генов в результате миграции зебувидного скота Индии в Африку. Использование генных частот позволяет вычислить генетические дистанции между породами и определить их эволюционную взаимосвязь. На рисунке 48 в качестве примера показаны эволюционные взаимосвязи между 14 породами скота.
По данным С. А. Петрушки (1970), частота аллеля p-LgA была в 2 раза выше у животных голландской и симментальской пород (0,514 и 0,436) в сравнении с бурой латвийской (0,210). Многие
|
|
|
 Скачать 5.3 Mb.
Скачать 5.3 Mb.