В. Н. Сайтаниди Рецензент членкорреспондент расхн в. Ф. Красота Петухов В. Л. и др. П31 Ветеринарная генетика В. Л. Петухов, А. И. Жигачев, Г. А. Назарова. 2е изд., перераб и доп. М. Колос, 1996. 384 с ил. Учебники
 Скачать 5.3 Mb. Скачать 5.3 Mb.
|

|
| Локус | Детерминанта | Класс Ig | Локализация |
| а | al, a2, аЗ | Все классы | V-область к-цепи |
| х, У | х32, уЗЗ | To же | Тоже |
| b | Ь4, Ь5, Ь6, Ь9 | » | С-область, к-цепи |
| с | с7, с21 | » | С-область, Х.-цепи |
| d | dll, dl2 | IgG | С-область, у-цепи |
| | | | (участок «талии» |
| | | | молекулы Ig) |
| с | е14, е15 | IgG | С-область, у-цепи (Fc- |
| | | | участок молекулы Ig) |
| f | fl59, f70, f71, f72, f73 | IgA | С-область, а-цепи |
| g | g74, g75, g76, g77 | IgA | То же |
| Локус не- | Msl, Ms2, Ms4, Ms5, Ms6 | IgM | С-область, ц-цепи |
| известен | | | |
| То же | n81, n82 | | То же |
Аллотипические детерминанты, указывающие аллотипичёскую группу (локус), обозначают буквой (a, b, e и т. д.), а аллели — цифрой (а1, Ь5, е14 и т. д.). При написании фенотипа аллотипы разных групп отделяют косой чертой, например а2аЗ/е14е15/Ь5, а генотип обозначают а2а3е14е15Ь5Ъ5.
234
Имеется еще две группы антигенных детерминант иммуноглобулинов: изотипы и идиотипы.
Изотипы — это антигенная специфичность, общая для всех особей одного вида. Пять классов иммуноглобулинов (А, М, С, Е, D) являются изотипами.
Идиотипы — это антигенные различия между антителами, принадлежащими к одному классу, субклассу и аллотипу у отдельных особей. Идиотипы одной особи определяются клонами клеток, синтезирующими антитела.
В зависимости от числа замен различают простые и сложные (комплексные) аллотипы. Простые аллотипы возникают при замене одной, двух аминокислот, сложные аллотипы — при замене многих аминокислот. Например, комплексные Ь4 и Ь9 аллотипы константного района к-цепи различаются по 35 % аминокислот.
Гены каждого вида цепи тесно сцеплены, поэтому их аллель-ные варианты наследуются как единое целое, т. е. аллогруппами (гаплотипами, группами сцепления). Например, три группы аллоти-'пов кролика (у, х, а) Ун-области и пять групп аллотипов (и, d, e, f, g) Сн-области наследуются аллогруппами. Ниже показаны алло-группы кролика, унаследованные потомками от отца и матери*
cL
к-цепь
Са2
Сн Сц Су Са1
aj_\ \n82\d"\e"\fn
? w"
I
\<>72i
\°2i \*" \"12
Известно, что в молекуле иммуноглобулина или только к-цепь, или только Х,-цепь сочетается с любым типом тяжелых цепей. В одной плазматической клетке вырабатываются антитела одной -специфичности. В гетерозиготных плазматических клетках работает только один из двух аллельных генов Vh- и Сн-цепей и один ген из двух легкой цепи — аллельное исключение (рис. 52). Поэтому фенотип клетки может не полностью соответствовать ее генотипу, несмотря на кодоминантное наследование. Однако в связи с наличием большого числа плазматических клеток в организме встречаются все типы антител в соответствии с его генотипом и любые комбинации материнских и отцовских аллотипов.
Разнообразие антител. Одна из главных и интересных проблем иммунологии — это объяснение природы происхождения громадного разнообразия антител. Ведь организмы в течение жизни могут встречаться с десятками тысяч антигенов и должны отличать «чужеродные» агенты от «своих». Но существует ли тогда
235
Bs
d"
Г

 ены V-обласпги t ^Гены С-обпасти
ены V-обласпги t ^Гены С-обпастиа* Транскрипция in
г
т
Антитело Ч-область С-область
Тяжелая цепь la
ШМ
Легкая цепь Ц
Рис. 52. Схема синтеза i лобулина. Генотип клетки — i мер адфльного исключения;
i клеткой тяжелых и легких i - -n/bJk i фенотип клетки — i а , d , d ,b,b— аллотипы иммуноглобулина кролика
огромное количество генов антител, передающихся из поколения в поколение?
Установлено, что разнообразие антител может быть обеспечено следующими факторами:
наличием ограниченного числа гаметных генов;
сборкой и экспрессией генов в соматических клетках из
ограниченного набора зародышевых сегментов;
неточностью аппарата сплайсинга РНК, когда соединяются
сегменты V, D и J;
соматическими гипермутациями генов антител.
Вам уже известно (см. рис. 50), что набор зародышевых генов для к-, Х-цепей и тяжелой цепи не столь велик: от 2 до 300 V-сегментов, 4 J-сегмента, около 20 D-сегментов и 1—4 С-сег-мента.
Количество вариантов сборки гена иммуноглобулина, например, легкой к-цепи равно произведению чисел V- и J—сегментов. Соединение V-, D- и J-сегментов может произойти более 10 тыс. способами. Таким образом, число вариантов формирования активного центра Ig (паратопа) участками легкой и тяжелой цепи превышает 10 млн.
Разнообразие антител может возникать вследствие нарушения соединения V-, D- и J-сегментов, а также вклинивания лишних нуклеотидов уже не в ДНК, а при сплайсинге про-иРНК. В результате образуется несколько измененная иРНК и при трансляции изменяется последовательность аминокислот в иммуноглобулине.
Значительный источник разнообразия антител — это повы-
236
шенная частота соматических мутаций в вариабельных доменах, особенно в гипервариабельных районах. Показано, что в V-райо-не Ig происходит три мутации на 30 делений В-лимфоцитов, т. е. на несколько порядков превышает обычную частоту мутаций. Предполагают наличие в В-клетках специальных ферментов, усиливающих темп мутирования.
Таким образом, при наличии различных вариантов сборки / зародышевых генов, неточности соединения сегментов, вставок пар нуклеотидов и соматических мутаций может возникнуть более 1 млрд типов антител.
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ ИММУННОГО ОТВЕТА
Иммунный ответ, или иммунологическая реактивность, — высокоспецифическая форма реакции организма на чужеродные вещества (антигены). При иммунном ответе происходят распознавание чужеродного агента и его элиминация. При введении антигена возникает первичный иммунный ответ — приблизительно через 2 дня в крови образуются антитела, титр которых возрастает, достигает максимума (к 4—6-му дню), а затем падает. Вторичный иммунный ответ возникает на повторное введение того же антигена и характеризуется более высоким и быстрым нарастанием титра антител. Подобная реакция более усиленного образования антител на повторное введение антигена, как вы знаете, называется иммунологической памятью, обусловлена наличием клеток иммунологической памяти и может сохраняться в течение многих месяцев и даже лет. Иммунный ответ зависит от генотипа организма:
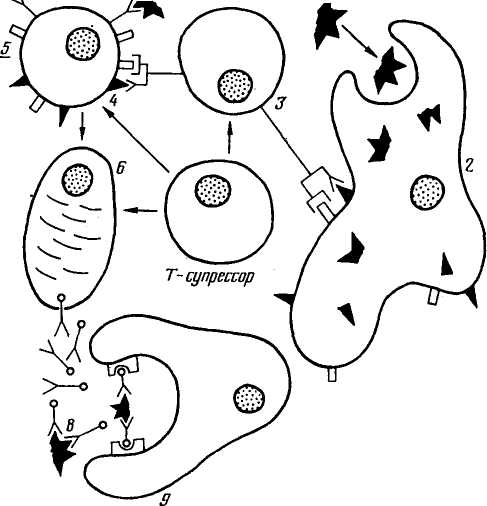
Иммунный ответ на инфекцию в общем виде показан на рисунке 53. Сначала антиген захватывается (I) представляющей клеткой (макрофагом). Внутри макрофага антиген перерабатывается (процессинг антигена) и попадает на его поверхность (2). Антиген на поверхности макрофага узнает активированный Т-хелпер (3). Т-супрессоры при иммунном ответе взаимодействуют с Т-хелперами и В-лимфоцитами. Т-хелпер активирует 1, В-лимфоцит, на поверхности которого находится процессирован-; ный антиген (4). В-лимфоциты на своей поверхности имеют ре-депторы-иммуноглобулины, которыми они узнают и связывают циркулирующий в организме антиген (5). Активированные В-лимфоциты пролиферируют и превращаются в клон плазматических клеток (б), а некоторые из их потомков становятся клетками памяти (7), обеспечивающими быстрый ответ на инфекцию в будущем. Клон плазматических клеток продуцирует антитела, которые связываются с антигеном, маркируют его (8), антигены узнаются и уничтожаются макрофагами (9).
При вирусной инфекции ДНК или РНК вируса попадает в клетку, а вирусные белки остаются на клеточной мембране
237

Вирус
Зараженная клетка
А Вирусный антиген
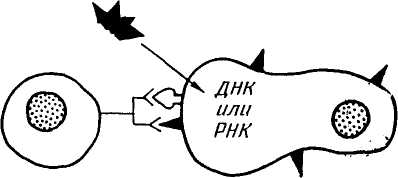 (рис. 54). Цитотоксические Т-клетки (Т-киллеры) своими рецепторами специфически узнают вирусные антигены только в комбинации с белком -главного комплекса гйстосовместимости (МНС) класса I, которые имеются почти на всех клетках организма. МНС класса I определяют иммунологическую индивидуальность клетки. В отличие от антител Т-рецепторы не узнают и не связывают антиген, если тот не находится вместе с белком МНС. После узнавания антигенов цитотоксические Т-клетки убивают зараженные вирусом клетки.
(рис. 54). Цитотоксические Т-клетки (Т-киллеры) своими рецепторами специфически узнают вирусные антигены только в комбинации с белком -главного комплекса гйстосовместимости (МНС) класса I, которые имеются почти на всех клетках организма. МНС класса I определяют иммунологическую индивидуальность клетки. В отличие от антител Т-рецепторы не узнают и не связывают антиген, если тот не находится вместе с белком МНС. После узнавания антигенов цитотоксические Т-клетки убивают зараженные вирусом клетки. Мутации любых локусов, обусловливающие разные звенья иммунной системы организма, в той или иной степени влияют на иммунный ответ. У кроликов, свиней, птицы, мышей и т. д. обнаружены индивидуальные и межлинейные различия по силе иммунного ответа.
Гены иммунного ответа (Ir-гены). При иммунизации инбред-ных мышей различными антигенами выявлены линии с сильным и слабым иммунным ответом. Гены, кодирующие иммунный ответ,<15ыли названы генами иммунного ответа (immune response).
Клеткп Активированный Активированный
Клон плазматических клеток
Антиген
ПроцессироВан-нып антиген
Иммуноглобулин* рецептор
у [
Еекретируемый иммуноглобулин
П Белок МНС (класс 11)
Рецептор г- хелпео
Клетка, пред-стабляшщая антиген (макрофаг) (А - клетки)
Макрофаг
Рис. S3. Иммунный ответ на инфекцию (по Тонегава, 198S)
п
 амятиВ-лимфоцит k Т-хелпер
амятиВ-лимфоцит k Т-хелпер238
Белок МНС класс!
Цитотоксическая Т-клетка (Т-киллер)
Рис. 54. Ответ на вирусную инфекцию (по Тонегава, 1985)
Высота иммунного ответа детерминирована многими генами иммунного ответа, обозначаемыми Ir-1, Ir-2 и т. д. У мышей известно более 20 1г-генов. Ir-гены контролируют ответ на тимуеза-висимые антигены и реализуют свое действие на уровне Т-лимфо-цитов. Контроль иммунного ответа осуществляется 1г-генами путем контроля синтеза la-белков (продукты генов 1-района МНС). Эти белки на мембране макрофага вступают в ассоциацию с молекулами процессированного (переработанного в макрофаге) антигена.
В I-районе главного комплекса гйстосовместимости мышей Н-2 существует три локуса Ir (Ir-1A, Ir-1B, Ir-lC). Кроме того, известны Ir-гены, расположенные вне Н-2 комплекса. Это локу-сы Ir-2, Ir-4, а также локус Ir, сцепленный с полом. Общее количество Ir-генов неизвестно. Ir-гены контролируют также клеточные реакции иммунитета. Во многих случаях иммунный ответ против антигенов наследуется полигенно.
Иммунизация свиней различными антигенами позволила также открыть гены иммунного ответа, которые имеют сходство с Ir-генами мышей. Иммунный ответ носит количественный характер, а Ir-гены сцеплены с главным комплексом гйстосовместимости свиней SLA.
Межлинейные и межпородные различия антителогенеза. Путем селекции можно усилить иммунный ответ на различные антиге-ны: При иммунизации лептоспирами мышей 12 инбредных линий и сублиний установлено, что титры антител у особей некоторых линий были в 2 раза выше, чем у других. При скре- Щивании контрастных линий обнаружено доминирование высокого иммунного ответа. Анализирующее скрещивание дало возможность сделать заключение о детерминации иммунного ответа более чем одной парой генов.
Обнаружены различия в титрах антител поросят разных Пород, вакцинированных модифицированной живой вирус-вак-Циной против псевдобешенства (табл. 36).
239
36. Средние титры антител (1дг) поросят разных пород после вакцинации против псевдобешенства (по Rothschild и др.)
Число поросят
Ig2 титра антител
П
 орода
орода96 75 132 152
4,29 ± 0,12 4,10 ±0,25 3,64 ± 0,11 3,46 ± 0,13
Йоркширская Гемпширская Дюрок Ландрас
К. В. Жучаев и С. П. Князев (1990—1993) установили у свиней значительную внутрипопуляционную дифференциацию по гуморальному иммунному ответу на сальмонеллы и эритроциты барана, по клеточной реакции на фитогемагглютинин: группы потомков разных хряков-производителей различались на 25—40 %.
Установлено, что у монозиготных двоен крупного рогатого скота наблюдается большее сходство в иммунном ответе на ряд антигенов, чем у дизиготных двоен.
Биози с соавт. (1973) в результате отбора инбредных мышей одной линии по высоте иммунного ответа на эритроциты барана в девятом поколении добились 30-кратных различий по титру антител между высоко- и низкореагирующими группами. Биози также проводил селекцию в течение 20 поколений и создал две линии мышей (АВ/А и AB/L), которые имели 200-кратные различия по титру антител против эритроцитов барана (1:10 000 и 1:40). Животные этих линий различались по высоте иммунного ответа ко многим, но не ко всем антигенам. Иммунологическая реактивность на некоторые антигены у животных этих линий оказалась одинаковой.
На основании вышесказанного можно сделать важный для селекционера вывод о том, что общей высокой иммунологической реактивности нет, а имеется конкретный иммунный ответ (Петров, 1983). Поэтому невозможно создать линии и породы животных, которые были бы в одинаковой мере резистентны ко всем болезням.
Очень важно для селекции то, что генетические различия между высокореагирующими и низкореагирующими особями на какие-то антигены сохраняются не только в течение жизни организмов, но даже после воздействия ионизирующей радиации — сильного иммунодепрессивного фактора. Так, воздействие сублетальных доз у-лучей на мышей, различающихся в 20 раз по продукции антител к лептоспирам, приводит к снижению титра антител в этих группах, но межлинейные различия сохраняются.
Можно сделать некоторые обобщения в отношении генов иммунного ответа:
1) Ir-гены определяют количество синтезируемых антител против определенных антигенов;
240
многие Ir-гены связаны с генами главного комплекса гис-
тосовместимости и отвечают за кооперацию Т- и В-лимфоцитов;
Ir-гены не сцеплены с локусами, кодирующими иммуног
лобулины;
Ir-гены высокоспецифичны. У организмов с одним и тем
же генотипом может быть высокий иммунный ответ против
одного антигена и низкий — против другого. Не обнаружены
гаплотипы, определяющие общую высокую или низкую иммуно
логическую реактивность;
генетически обусловленные различия в высоте иммунного
ответа сохраняются в разные возрастные периоды;
между генами, контролирующими высокий или низкий
иммунный ответ против различных антигенов, в основном не
существует никакой связи;
эффект генов, отвечающих за иммунологическую реактив
ность, реализуется на уровне популяции лимфоидных клеток.