В. Н. Сайтаниди Рецензент членкорреспондент расхн в. Ф. Красота Петухов В. Л. и др. П31 Ветеринарная генетика В. Л. Петухов, А. И. Жигачев, Г. А. Назарова. 2е изд., перераб и доп. М. Колос, 1996. 384 с ил. Учебники
 Скачать 5.3 Mb. Скачать 5.3 Mb.
|

|
225 Герефордская 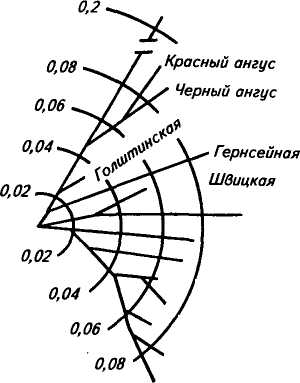 Исландская Джерсейская Исландская Джерсейская Шороле. Retinto \ Испанские Delidia J п°Р°°ы Иберийский Mertobenga\ Португальские Alentejana JпоРоды Американская длиннорогая Рис. 48. Эволюционные взаимосвязи между 14 породами скота, вычисленные на основании генетического расстояния по 14 локусам. Филогенетическое древо изображено в полярных координатах; расстояние оценивалось способом наименьших квадратов радиальной длины каждого сегмента (по Kidd и др.,. 1980) европейские породы имеют очень низкую частоту типов транс-феррина Tf8 и Tf*\ Использование полиморфных систем белков вместе с группами крови повышает точность определения происхождения животных. Так, по группам крови отцовство можно установить в 81 % случаев, а дополнительные анализы только типов транс-феррина повышают точность до 90 %. По данным В. В. Пилько, Ю. О. Шапиро и А. С. Гурьяновой (1970), в четырех хозяйствах Белоруссии у коров бурой латвийской и костромской пород с TfDD удой был выше на 256— 270 кг, чем у животных с другими генотипами. Л. С. Жеброский и др. (1979) на большом материале показали, что ген к-СпА во всех стадах связан с повышенной молочностью. Таким же эффектом обладает аллель P-LgA, но в то же время он снижал содержание жира в молоке коров черно-пестрой породы. Данные по красной датской породе свидетельствуют о том, что только 3 % генетической изменчивости в содержании жира и 5 % в молочности обусловлены различиями по группам крови. Видимо, есть большая вероятность установления более тесной корреляции генетических полиморфных систем с резистентное -тью к некоторым заболеваниям вследствие менее сложной их генетической детерминации, чем признаков продуктивности. 226 Анализ полиморфизма яичного белка овоглобулинового локу-са G3 показал, что куры с типом АВ более устойчивы к пуллоро-зу—тифу. Восприимчивость к этому заболеванию кур породы леггорн была связана с аллелем G^S, а породы корниш — с алле-лем GB3. В Австралии, а потом в Кении у породы овец ромни-марш с типом гемоглобина НЬА найдена более высокая резистентность к гемонхозу (заболевание, вызываемое нематодами, паразитирующими в сычуге), чем у животных с гемоглобином типов НЬВ и НЬАВ. Однако имеются сведения и об отсутствии связи типов гемоглобина у местных флоридских овец с невосприимчивостью к гемонхозу. Устойчивость овец к лептоспирозу связана с гетерозиготнос-тью по гемоглобиновому локусу (НЬАВ), тогда как особи с типом А или В более восприимчивы. Эта инфекционная болезнь проявляется у животных кратковременной лихорадкой, желтухой, гемоглобинурией, абортами и другими признаками. У свиней найдена ассоциация лептоспироза с аллелем амилазы АпА Уровень антител к лептоспирозу увеличивался у животных с этим аллелем, а с аллелем Anv8 — уменьшался. У свиней установлена связь типов фермента фосфогексоизо-мераза (PHI) с синдромом злокачественной гипертермии (MHS). Коэффициент корреляции между чувствительностью к MHS и генотипом РН1В/РН1В равен 0,19, а относительный риск возникновения MHS у особей этого генотипа по отношению к имеющим его животным был 18,8. Изучение новых биохимических полиморфных систем позволит глубже понять динамику генотипической изменчивости в популяциях и механизмы поддержания этой изменчивости, изменение генетической структуры популяций при селекции, планирование и контроль с их помощью селекционного процесса. Можно рассчитывать на выявление более однозначных связей отдельных аллелей или их совокупности с резистентностью к болезням, признакам продуктивности и использовать полиморфные системы как генетические маркеры в селекции. Контрольные вопросы. 1. Что такое генетическая система групп крови, тип крови, феногруппа? 2. В чем заключаются особенности наследования групп Крови? 3. Как определяются группы крови у животных? 4. Какие теоретические предпосылки лежат в основе связи групп крови с продуктивностью и устойчивостью к болезням? 5. Какое значение группы крови имеют для практики? 6. Почему возникает гемолитическая болезнь новорожденных? Какие методы профилактики этой болезни вы знаете? 7. Что лежит в основе генетического полиморфизма? 8. Какое значение для практики имеет биохимический полиморфизм? 227 Г л а в а 14 ГЕНЕТИЧЕСКИЕ ОСНОВЫ ИММУНИТЕТА Иммунитет — невосприимчивость организма к инфекционным агентам и генетически чужеродным веществам антигенной природы. По Р. В. Петрову (1983), иммунитет — способ защиты организма от живых тел и веществ, несущих на себе признаки генетической чужеродности. Главная функция иммунитета — иммунологический надзор за внутренним постоянством (гомеостазом) организма. Следствие* этой функции — распознование, а потом специфическое блокирование, нейтрализация или уничтожение генетически чужеродных веществ (бактерий, вирусов, раковых клеток и т. д.). За сохранение генетически обусловленной биологической индивидуальности отвечает иммунная система организма — совокупность всех лимфоидных органов и скоплений лимфоидных клеток. Она состоит из центральных и периферических органов. Центральные органы иммунной системы включают тимус, сумку Фабри-ция (у птиц) и ее аналог у млекопитающих, костный мозг, пейеровы бляшки и миндалины. К периферическим органам относят лимфатические узлы, селезенку и кровь. Иммунная система и ее главные исполнители — лимфоциты обеспечивают специфическую реакцию организма на чужеродные антигены. Кроме этой системы существуют механизмы, обеспечивающие первичную ступень неспецифической сопротивляемости организма паразитам (бактериям, вирусам, гельминтам и т. д.). К неспецифическим факторам защиты относят кожные и слизистые покровы, фагоциты (нейтрофилы, тканевые макрофаги), естественные иммуноглобулины, систему комплемента (включающая около 20 белков), интерферон, лизоцим, пропердин, лактоферрин и т. д. Неспецифические факторы защиты действуют в широком спектре, хотя ряд из них может быть в большей или меньшей степени направлен против некоторых групп микроорганизмов. Интерферон обладает противовирусным действием, пропердин характеризуется выраженным антимикробным свойством, комплемент осуществляет бактериологическое действие. В то же время фагоциты и комплемент участвуют и в специфических реакциях. Фагоциты, кооперируясь с Т- и В-лимфоци-тами, принимают участие в иммунном ответе. 228 Большинство защитных механизмов организма находится под генетическим контролем. Видимо, содержание лизоцима комплемента, пропердина и других веществ наследуется полигенно. На генетическое разнообразие по этим признакам указывают межпородные, межлинейные и межсемейные различия. Имеются генетические дефекты, ведущие к потере способности нейтрофи-лов к фагоцитозу. Один из них впервые открыт у человека и назван синдромом Чедиака—Хигаши, а сейчас он известен и у крупного рогатого скота (особенно герефордской породы), норок, бизонов, мышей и др. Синдром характеризуется наличием в цитоплазме лейкоцитов больших гранул. У крупного рогатого скота это приводит к частичному альбинизму, светобоязни и чувствительности к инфекциям. У алеутских голубых норок известна алеутская болезнь, вызываемая вирусом, при которой наблюдаются нарушения, как и при синдроме Чедиака—Хигаши. Синдром у скота и норок наследуется как аутосомно-рецессив-ный признак. Заболевают гомозиготные особи (аа). Было также обнаружено у человека и собак генетически детерминируемое снижение числа нейтрофилов — синдром циклическая нейтропения. Эта болезнь встречается у колли с серым окрасом, которые погибают до полового созревания. Рецессивный ген серой окраски у колли обладает и плейотропным эффектом ri отношении этого летального дефекта. Клеточная и гуморальная системы иммунитета. Стволовые лимфоидные клетки, мигрировавшие в тимус, превращаются в Т-лимфоциты (Т-клетки), которые ответственны за клеточную форму иммунного ответа, а сформированные в сумке Фаб-риция (у птиц) или ее аналоге у млекопитающих становятся В-лимфоцитами (В-клетки), которые ответственны за реализацию гуморального иммунного ответа. Т- и В-системы лимфоцитов состоят из субпопуляций клеток. Так, Т-клетки образуют субпопуляцию Т-хелперов, Т-супрессоров, Т-киллеров. Первые из них способствуют превращению В-лимфоцитов в плазматические клетки, вторые блокируют антителообразование В-лимфоцитами и участвуют в становлении и поддержании иммунологической толерантности, третьи разрушают клетки чужеродных трансплантантов и злокачественные клетки. Многообразие иммунологических реакций является следствием кооперации Т- и В-лимфоцитов и макрофагов, в результате чего образуются антитела (иммуноглобулины). Синтез антител осуществляется вплазматическихклетках, происходящих из В-лимфоцитов. Одна клетка после стимуляции антигена за 1 с секретирует около 2 тыс. молекул одной специфичности и определенного класса. На поверхности В- и Т-лимфоцитов имеются рецепторы им-муноглобулиновой природы, причем на В-лимфоцитах их в де- 229 сятки и сотни раз больше. Рецепторы — это макромолекулярные структуры клеточной поверхности, с помощью которых клетки узнают антигены. Поэтому проблема специфических рецепторов — одна из центральных в иммунологии, так как благодаря этому происходит распознавание генетически «своего» и «чужого». Синтез и специфичность рецепторов контролируются генетически. В-система в большей степени ответственна за иммунитет при многих бактериальных инфекциях, антитоксический иммунитет, анафилаксию, аллергию немедленного типа, некоторые аутоиммунные заболевания (красная волчанка и др.). Т-система играет главную роль в иммунитете против большинства вирусных инфекций, туберкулеза, бруцеллеза, туляремии, в трансплантационном и противоопухолевом иммунитете, аллергии замедленного типа, ряде иммунопатологии и др. СТРУКТУРА ИММУНОГЛОБУЛИНОВ Иммуноглобулины — семейство белков, специфически реагирующих с антигеном, который индуцировал их образование. Термины «антитело» и «иммуноглобулин» — синонимы. У большинства млекопитающих иммуноглобулины разделяют на 5 классов: IgG, IgA, IgM, IgD, IgE. Например, у взрослых свиней есть 3 типа сывороточных иммуноглобулинов: IgG, IgA, IgM и 2 подкласса IgG (19S и 7S), которые по физико-химическим свойствам аналогичны соответствующим Ig человека. Кроме сыворотки крови иммуноглобулины находятся в молозиве, молоке, слюне, секретах кишечника и т. д. У всех видов животных и человека молекулы Ig построены из двух длинных тяжелых (Н) и двух коротких легких (L) полипептидных цепей, соединенных дисульфидными мостиками. Для всех классов Ig легкие цепи являются общими, а тяжелые цепи каждого из 5 классов отличаются по антигенным, иммунологическим и химическим особенностям. Тяжелые цепи иммуноглобулинов обозначают буквами греческого алфавита. IgD IgE 8 (дельта) е (эпсилон) Иммуноглобулин Тяжелые цепи IgG IgA IgM у (гамма) а (альфа) ц (мю) легких цепях, которые обозначаются Vh, ChI, Сн2, СнЗ и Vl.Cl (рис. 49). Домены Vh и Vl выполняют функцию связывания антигена. Иммуноглобулин G расщепляется ферментом папаи-ном на три фрагмента: два Fab-фрагмента связывают антиген и один Fc-фрагмент ответствен за связывание белка комплемента, реакцию с макрофагами и др. Реакция антиген — антитело. Основной момент в процессе иммунного ответа — узнавание антителом химического маркера, характерного «чужому» веществу в отличие от «своего». Поэтому главная биологическая функция антител — их способность вступать в специфическую и быструю реакцию с антигеном, в результате чего образуется комплекс антиген — антитело (иммунный комплекс). Иммунный комплекс образуется в результате связывания активного центра антитела (п а р а т о п а) с детерминантами антигена (э п и т о п а). Эти взаимодействия могут проявляться в виде реакций агглютинации, преципитации, лизиса, нейтрализации и др. Антитела могут усиливать фагоцитарную активность макрофагов (о п с о н и з а ц и я). Специфичность иммунитета проявляется в том, что антитела действуют только на тот антиген, под влиянием которого они образовались. Организм, имеющий антитела, может оставаться в течение различного времени иммунным 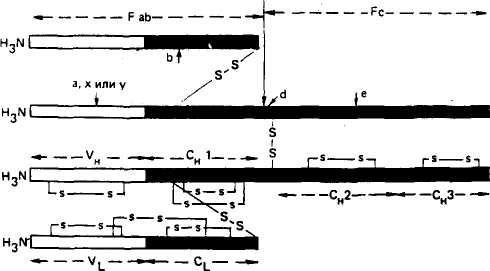 соон cooh 120 280 40 360 200 440 Легкие цепи подразделяют на два типа: каппа (к-тип) и лямбда (Х-тип). N-концевые участки тяжелых и легких цепей состоят из вариабельных (V) областей, а С-концевые участки тяжелых и легких цепей являются константными (С). Цепи Ig представлены не в виде прямых нитей, а образуют петли, возникающие при соединении дисульфидными мостиками аминокислотных остатков внутри цепи. Каждая петля называется доменом. В молекуле Ig кролика 12 доменов — по 4 на тяжелых и по 2 на 230 НОМЕР АМИНОКИСЛОТНОГО ОСТАТКА Рис. 49. Схема молекулы IgG кролика: £ —легкие и Я —тяжелые цепи; V— вариабельные области легких и тяжелых цепей (светлые); С — константные области зачернены; s — s — дисульфидные связи; Vl и Cl — домены, составляющие вариабельные и константные участки легких цепей, Ун, Сн1, Сн2, СнЗ— .домены, составляющие вариабельные и константные участки тяжелых цепей; Fab (два) и Fc (один) — фрагменты, образующиеся при расщеплении папаином; а, х, у, b, d, e — локализация известных аллотипических групп 231 Гены легкой А- цепи \\ V " J С J Гены легкой Н- цепи J С J С Интрон 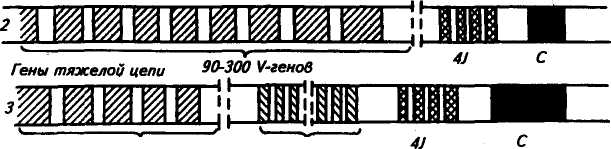 против антигена. Следовательно, гуморальный, как и клеточный, иммунитет обладает иммунологической памятью — способностью при повторном контакте с антигеном узнавать и отвечать на него иммунологической реакцией. против антигена. Следовательно, гуморальный, как и клеточный, иммунитет обладает иммунологической памятью — способностью при повторном контакте с антигеном узнавать и отвечать на него иммунологической реакцией. В определенных условиях возникает повышенная чувствительность организма (гиперчувствительность) на повторное попадание данного антигена. Она проявляется в виде аллергии немедленного типа (астма, анафилаксия), т. е. патологической повышенной реакции на антиген, который у нормальных особей не вызывает болезненных явлений. Существует и повышенная чувствительность замедленного типа (при туберкулезе, бруцеллезе) — это клеточно опосредованный иммунологический способ повышенного реагирования на чужеродные вещества. Антитела участвуют в первой форме иммунного реагирования — гуморальном иммунитете. Вторая форма больше связана с клеточным иммунным ответом в виде гиперчувствительности замедленного типа. Прфтолагают, что иммуноглобулин может иметь несколько антигенсвязывающих центров, комплементарных нескольким отличным по структуре антигенам, а многие типы антител комплементарны к одной антигенной детерминанте. Антитело способно функционировать и как антиген. Высокоспецифическая популяция антител может состоять из иммуноглобулинов разной специфичности. Очень небольшие изменения в первичной структуре антител могут вызвать различия в их специфичности. ГЕНЕТИКА ИММУНОГЛОБУЛИНОВ Гены иммуноглобулинов аутосомны, кодоминантны. Структурное разнообразие иммуноглобулинов определяется последовательностью аминокислот. В биологии существовал принцип «один ген — одна полипептидная цепь», а также утверждалась неизменность генома в онтогенезе организма. Однако в случае Ig видно, что несколько генов кодируют одну полипептидную цепь. Иммуноглобулины контролируются тремя семействами генов, расположенными в разных хромосомах. Одно семейство кодирует синтез всех классов тяжелых цепей (Н), другое — синтез легких к-цепей, третье — синтез легких 1-цепей. У мыши вариабельная область легкой А,-цепи кодируется двумя V-генами, а константная область — четырьмя С-генами (рис. 50). V- и С-гены называются также V- и С-сегментами. Перед каждым С-геном находится короткий отрезок ДНК, который называется J-сегментом. Любой V-ген может соединиться с любой парой J-сегмент — С-ген. Следовательно, легкая А,-цепь кодируется сегментами'V, J, С. В эмбриональном геноме эти 232 20D 100-200 V-генов Рис. 50. Гены Ig шали (1 — легкая Х-цеш»; 2 — легкая к-цепь; 3 — тяжелая цель; V—ген (сегмент); J—сегмент; D—сегмент; С—ген (сегмент) сегменты удалены друг от друга, а в зрелых лимфоцитах в результате соматической рекомбинации они сближаются. Для легкой к-цепи имеется несколько сот V-генов, четыре J-сегмента и один С-ген (см. рис. 50). В зрелом лимфоците Я,-цепь кодируется сегментами V, J и С (по одному из многих сегментов. V и J). Для тяжелых цепей всех типов существует 100—500 V-генов, 20 D-сегментов, 4 J-сегмента и несколько тесно сцепленных С-генов. В плазматической клетке тяжелая цепь кодируется сегментами V, D, J, С (см. рис. 50). Сбор генов Ig осуществляется в два этапа. В результате соматической рекомбинации соединяются V- и J-сегменты легкой цепи или V-, D- и J-сегменты тяжелой цепи. Часть V- и J-cer-ментов при этом выбрасывается. Затем происходит транскрипция, образуется пре-иРНК. При сплайсинге про-иРНК (удаляются интроны и соединяются экзоны) вырезаются интроны и V-, J-, С-сегменты располагаются рядом. В цитоплазме происходит трансляция и образуется легкая к-цепь (рис. 51). Такой же процесс осуществляется и при образовании тяжелых цепей Ig. При иммунном ответе предшественники зрелых лимфоцитов сначала образуют филогенетически самый древний IgM, а потом в лимфоцитах происходит переключение на синтез Ig других классов. Аллотипы иммуноглобулинов. Это генетически детерминированные варианты белков (в данном случае иммуноглобулинов), по которым особи внутри вида отличаются друг от друга. Аллотипы представляют отдельные варианты генов иммуноглобулинов и обусловливают их генетический полиморфизм. Каждая молекула иммуноглобулина имеет не один, а несколько аллоти-пических детерминант (или аллотипических маркеров). Из сель- 233 Ш: 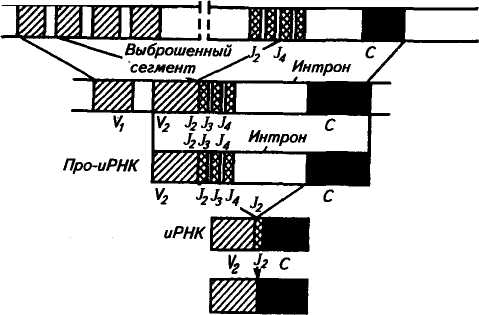 |