В. Н. Сайтаниди Рецензент членкорреспондент расхн в. Ф. Красота Петухов В. Л. и др. П31 Ветеринарная генетика В. Л. Петухов, А. И. Жигачев, Г. А. Назарова. 2е изд., перераб и доп. М. Колос, 1996. 384 с ил. Учебники
 Скачать 5.3 Mb. Скачать 5.3 Mb.
|

34,5 + (5-1)7,91 " 66,14
j "о "оо "ю чо Хп "н- чо Хл "о "§->»*■ ,».*.».ui o," S."1 и," G.» S.» 5 ^ и 5* soooevooMMmwK.» wo-n^^u^o\;., о;., ом., ч;,-„ ^Д Д S yj^4UJ^iLOyiiU>jJtUJWlU>O4^.^J^00*fcO04^4OLriQ(-h^CTs™4O р S • и "о "- |
 Й 2 5 V V "u>
Й 2 5 V V "u> 


| | |
| И о | II |
| | en |
| | VO |
| | |
"Зо
= 0,117,
11,7
2,4-0,19 _ 2,21
Таким образом, различия в устойчивости дочерей к бруцеллезу на 11,7 % зависят от генотипа их отцов и на 88,3 % —от других причин.
Контрольные вопросы. 1. Какие вы знаете виды изменчивости? 2. Что такое биометрия? 3. Какие статистические показатели характеризуют совокупность? 4. Какие показатели характеризуют изменчивость? 5. Как можно оценить взаимосвязь между признаками? 6. Как оценить достоверность разности между средними арифметическими двух выборочных совокупностей? 7. Для чего используется дисперсионный анализ?
154

 Г л а в а 10 МУТАЦИОННАЯ ИЗМЕНЧИВОСТЬ
Г л а в а 10 МУТАЦИОННАЯ ИЗМЕНЧИВОСТЬВ предыдущих главах отмечено одно из основных свойств кариотипа, ДНК и ее участков (генов) — сохранять постоянство внешнего и внутреннего строения. Морфофункциональная устойчивость генетического материала обеспечивает передачу всей совокупности наследственных признаков каждой особи последующим поколениям и является основой для сохранения видовых признаков на протяжении многих сотен лет. Однако такая стабильность относительна. В силу действия внутренних и внешних факторов в генетическом материале возникают изменения — мутации, определяющие мутационную изменчивость.
Мутациями называют стойкие изменения в структуре ДНК и кариотипе. Этот термин впервые предложил ботаник Гуго де Фриз для обозначения внезапно возникающих наследуемых изменений у растений. Большой вклад в развитие теории мутаций внесли отечественные ученые С. И. Коржинский, Н. В. Тимофеев-Ресовский А. С. Серебровский, Н. П. Дубинин, М. Е. Лобашов и др.
Мутации у животных происходят постоянно с определенной частотой и скоростью. Процесс образования их получил название мутагенеза. Мутации, возникающие в естественных условиях, называют спонтанными, искусственно вызванные — индуцированными. Те и другие могут возникать как в генеративных, так и в соматических клетках. Мутации, возникающие в половых клетках, передаются в последующие поколения. Соматические мутации не наследуются. Они влияют только на признаки самого мутантного животного.
КЛАССИФИКАЦИЯ МУТАЦИЙ
Основные типы мутаций — изменения в числе или структуре хромосом — хромосомные мутации, в структуре ДНК —генные (точковые) мутации.
Хромосомные и генные мутации, как правило, вызывают у животных нарушения жизнеспособности, плодовитости, снижение устойчивости к болезням, продуктивности и другие вредные последствия. Это связано с тем, что они приводят к нарушению процессов деления клеток, нормального распределения хромосом между ними, изменяют ход синтеза белков, ферментов и т. д.
156
ХРОМОСОМНЫЕ МУТАЦИИ
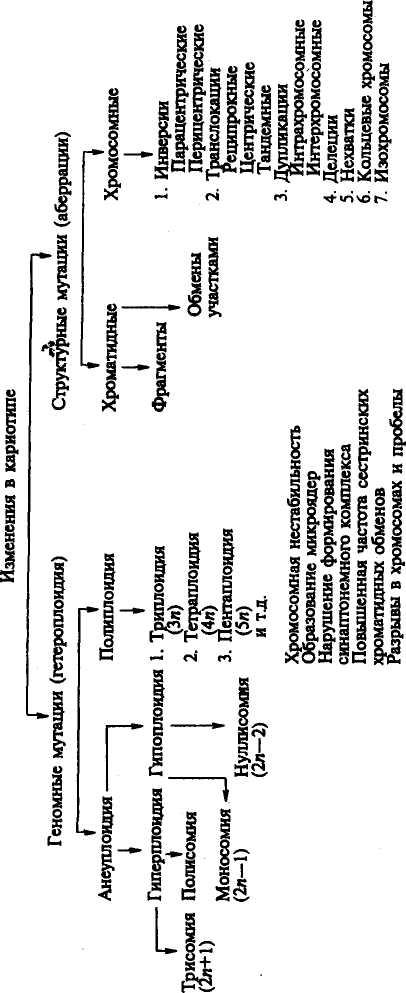
Изменения кариотипа могут быть количественными, структурными и одновременно теми и другими. Рассмотрим отдельные формы изменения хромосом (см. схему).
Числовые мутации кариотипа. Эта группа мутаций связана с изменением числа хромосом в кариотипе. Количественные изменения в хромосомном составе клеток называют геномными мутациями. Они подразделяются на гетероплоидию, анеуплоидию, полиплоидию.
Гетероплоидия обозначает общее изменение числа хромосом по отношению к диплоидному полному набору.
Об анеуплоидии говорят в тех случаях, когда число хромосом в клетке увеличено на одну (трисомия) или более (полисемия) или уменьшено на одну (моносомия). Употребляют также термины «гиперплоидия» и «гипоплоидия». Первый из них означает увеличенное число хромосом в клетке, а второй — уменьшенное.
Полиплоидией называют увеличение числа полных хромосомных наборов в четное или нечетное число раз. Полиплоидные клетки могут быть триплоидными, тетраплоидными, пентаплоид-ными, гексаплоидными и т. д.
Структурные мутации хромосом. Эта группа мутаций связана с изменением формы, размеров хромосом, порядка расположения генов (изменение групп сцепления), утратой или добавкой отдельных фрагментов и т. д. Изменения структуры одной или нескольких хромосом называют хромосомными мутациями. Установлено несколько типов структурных мутаций хромосом.
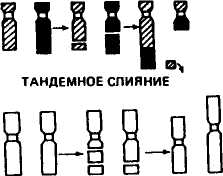
Транслокации — перемещения отдельных фрагментов хромосом из одного участка в другой, обмены фрагментами между разными хромосомами, слияния хромосом. При взаимных обменах фрагментами между гомологичными или негомологичными хромосомами возникают транслокации, называемые реципрокными. Если целое плечо одной хромосомы присоединяется к концам другой хромосомы, такой тип транслокаций называют тандемным. Слияние двух акроцентрических хромосом в области центромер формирует транслокацию робертсоновского типа и образование мета-и субметацентрических хромосом. При этом обнаруживается элиминация блоков прицентромерного гетерохроматина.
Инверсии — внутрихромосомные аберрации, при которых фрагменты хромосом разворачиваются на 180°. Различают пери-и парацентрические инверсии. Если перевернутый фрагмент содержит центромеру, инверсия называется перицентрической.
Делеции — потеря срединного фрагмента хромосомы, в результате чего она укорачивается.
Нехватки — потеря концевого фрагмента хромосомы.
Дупликация — удвоение фрагмента одной хромосомы (интра-
157
158
хромосомные дупликации) или разных хромосом (интерхромосомные дупликации).
Кольцевые хромосомы формируются при наличии двух концевых разрывов (нехваток).
РЕЦИПРОКНАЯ ТРАНСЛОКАЦИЯ
ПАРАЦЕНТРИ- ПЕРИЦЕНТРЫ- ПЕРЕМЕЩЕНИЯ
ЧЕСКАЯ ЧЕСКАЯ
ИНВЕРСИИ
8
ВСТАВКА
ЦЕНТРИЧЕСКОЕ СЛИЯНИЕ
S
ДЕЛЕЦИЯ
КОЛЬЦЕВАЯ ХРОМОСОМА
ДУПЛИКАЦИЯ
Изохромосомы возникают, если в противоположность нормальному делению хроматид в длину происходит горизонтальное (поперечное) деление хромосомы в центромере с последующим слиянием гомологичных плеч в новую хромосому — изохромосому. Ее проксимальные и дистальные участки идентичны по строению и составу генов. В зависимости от того, сколько хроматид изменено (одна или две), структурные аномалии подразделяются на хромосомные и хро-матидные. На рисунке 34 приведены схемы образования различных типов структурных изменений хромосом или аберраций.
ОБРАЗОВАНИЕ ИЗОХРОМОСОМ
НОРМАЛЬНОЕ РАСХОЖДЕНИЕ ХРОМАТИД
Рис. 34. Схема образования различных типов структурных аберраций хромосом
(по Харе, 1978)
159
НЕРАСХОЖДЕНИЕ ХРОМОСОМ В МЕЙОЗЕ
НЕ РАСХОЖДЕНИЕ ПРИ ПЕРВОМ МЕЙО-ТИЧЕСКОМ ДЕЛЕНИИ
ОДИН ИЗ ДВУХ
НЕРАСХОЖДЕНИЕ ПРИ ВТОРОМ МЕЙОТИЧЕСКОМ ДЕЛЕНИИ
ООЦИТЫ I ПОРЯДКА
ООЦИТЫ II ПОРЯДКА
оотиды
МЕХАНИЗМ ОБРАЗОВАНИЯ ЧИСЛОВЫХ И СТРУКТУРНЫХ АНОМАЛИЙ
КАРИОТИПА
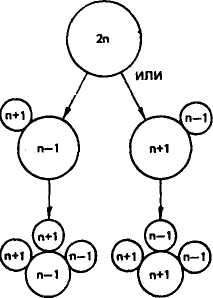
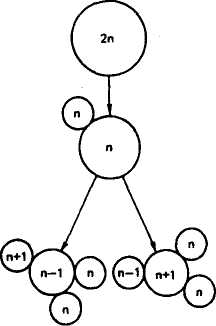
В основе всех числовых аномалий кариотипа лежит нерасхождение хромосом в мейозе. Чаще это происходит у особей женского пола. Так, трисомия и моносомия возникают вследствие нерасхождения хроматид во время мейотического деления. В результате в одной дочерней клетке появляется лишняя хромосома, в другой ее недостает. Полисемия, как и моносомия, возникает, очевидно, из-за нерасхождения во время мейоза или раннего дробления. В большинстве случаев трисомные и моносомные эмбрионы появляются в результате новых мутаций в половых клетках родителей. Наиболее часто у животных обнаруживают трисомию и моносомию половых хромосом. Вероятность нерасхождения хромосом увеличивается с возрастом животных.
Полиплоидия может возникать в результате ошибок при мейозе^ши во время оплодотворения. Так, при делении хромосом на хроматиды без деления цитоплазмы в клетке может образовываться тетраплоидный набор. Такие клетки могут возникать при слиянии двух соматических клеток. У животных обнаруживали диплоидные гаметы, которые формировались в результате нерасхождения хромосом в мейозе. Эти случаи полиплоидии связаны с нарушением образования веретена или тянущих нитей в анафазе.
Полиплоидия может быть следствием оплодотворения одной яйцеклетки с гаплоидным набором хромосом двумя или более спермиями (полиспермия или полиандрия). При оплодотворении одной яйцеклетки двумя спермиями формируется тршшоидный эмбрион. Триплоидный набор может возникнуть тогда, когда в процессе оплодотворения наряду с яйцеклеткой участвует и второе полярное тело (рис. 35). В других случаях полиплоидные индивидуумы возникают при задержке первого деления дробления зиготы.
Тетраплоидия может возникать благодаря подавлению первого дробления диплоидной зиготы, когда хромосомы дуплицируются и делятся, но остаются в той же клетке.
Предрасполагающие к появлению полиплоидии факторы: старение гамет при задержке овуляции, длительность хранения спермы, задержка оплодотворения самок. Слияния клеток могут индуцироваться вирусами. На образование веретена деления (разрыва нитей) могут влиять лекарственные препараты.
Мозаицизм и миксоплоидия, а также химеризм относятся к категории соматических мутаций. Мозаицизм — присутствие в организме клеток (точнее, клонов) разного генотипа, что может привести к возникновению в процессе соматического развития клеточных популяций с отличающимся генотипом. Частным случаем мозаицизма является гинандроморфизм и мозаицизм по
160
Рис. 35. Механизм образования трисомин и моносомин в мейозе (по Харе, 1978)
группам крови, белкам и др. Миксоплоидия, полисоматия — форма клеточного мозаицизма — наличие у одной особи клеток с различным уровнем плоидности (три-, тетра-, пента-, гекса-плоидные и т. д.). Миксоплоидные клетки возникают в результате нарушения митоза во время раннего дробления при делении зиготы (нерасхождение хромосом). Химеризм возникает в результате обмена клетками крови между плодами при двух или более плодной беременности, в случае слияния бластоцист или зигот. В частности, диплоидно-триплоидные химеры могут формироваться при слиянии второго полярного тела с одним из бластомеров на первой стадии дробления.
Структурные изменения в кариотипе {аберрации) возникают как результат спонтанной или индуцированной ломки, разрывов и последующих воссоединений хромосом новым способом. Если разрывы затрагивают обе хроматиды, аберрации называют хромосомными, если только одну — хроматидными. Если в результате структурной перестройки нет потери или прибавления генетического материала, аберрации считаются сбалансированными. К ним относятся в основном транслокации, инверсии. Несбалансированные аберрации представлены главным образом деле-циями, дупликациями, изохромосомами и кольцевыми хромосомами. После разрывов хромосом или хроматид оторвавшиеся фрагменты обычно утрачивают. Это приводит к потере части генов (делеции и нехватки) или дополнению хромосом фрагментами (дупликации), что связано с появлением избыточного гене-
161
■xz
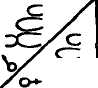 тического материала в клетках. Следствием делеций и нехваток может быть отсутствие генетического контроля для самых разных признаков организма. Рецессивные гены, содержащиеся в тех же локусах нормальной гомологичной хромосомы, проявляют свои эффект в одинарной дозе. Следует иметь в виду, что животные, гетерозиготные по сбалансированным транслокациям, могут производить генетически несбалансированные гаметы, которые, в свою очередь, могут формировать несбалансированные зиготы, отмирающие на ранних стадиях эмбриогенеза (рис. 36).
тического материала в клетках. Следствием делеций и нехваток может быть отсутствие генетического контроля для самых разных признаков организма. Рецессивные гены, содержащиеся в тех же локусах нормальной гомологичной хромосомы, проявляют свои эффект в одинарной дозе. Следует иметь в виду, что животные, гетерозиготные по сбалансированным транслокациям, могут производить генетически несбалансированные гаметы, которые, в свою очередь, могут формировать несбалансированные зиготы, отмирающие на ранних стадиях эмбриогенеза (рис. 36). В связи с широким распространением центрических слиянии (транслокаций Робертсона) у животных ученые активно изучают механизмы их образования. Полагают, что робертсоновские транслокации представляют собой частный вариант теломерного слияния, вовлекающего теломерные концы двух различных тело-центрических (акроцентрических) хромосом, у которых вследствие задержки репликации ДНК в области палиндрома либо точ-ковых мутаций в этой области рестрикционные эндонуклеазы утрачивают способность узнавать место рестрикции, что ведет к сохранению прочного соединения между хромосомами. При этом возникает метацентрическая или субметацентрическая хромосома, несущая две центрометры, которые либо функционируют как одно целое, либо одна центромера инактивируется.
Реципрокные транслокации у родителей — одна из основных причин возникновения трисомий и моносомий у эмбрионов. Частичные трисомий могут быть связаны как с транслокациями и инверсиями у родителей, так и с новыми мутациями-спорадические транслокации, дупликации и т. д. Необходимо отметить, что в данном случае терминами «частичные трисомий» и «моносомий», обозначаются потери части хромосом, т е делеций и нехватки. При транслокациях число плеч хромосом в клетках остается таким же, однако в измененных хромосомах образуются новые группы сцепления между генами, что нарушает процессы конъюгации гомологичных хромосом в мейозе и является причиной формирования несбалансированных зигот.
Хромосомная нестабильность. В процессе ци-тогенетического анализа можно выделить животных, не имеющих в кариотипе каких-либо изменений, и особей, у которых находят разрывы и пробелы хромосом, полиплоидные клетки, другие структурные и числовые аберрации. По специальным методикам у одних индивидуумов обнаруживают нарушения формирования синаптонемного комплекса в мейозе, повышенную частоту сестринских хроматидных обменов и высокий процент клеток с микроядрами. Повышенная частота числовых и структурных аномалий хромосом, наблюдаемая у отдельных особей, определяется термином «хромосомная нестабильность».
/ишхмлгя
iq
о!