ответы на физиологию. 1. Современные представления о строении и функции мембран
 Скачать 0.69 Mb. Скачать 0.69 Mb.
|

|
24. Формы постсинаптического торможения в ЦНС. Поскольку в нервных сетях включено огромное число интернейронов, они могут, передавая нервные импульсы другим нейронам центра, реали¬зовывать как их возбуждение, так и торможение. Различают следующие типы тормозных процессов в нейронных сетях: • 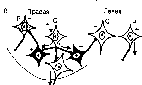 реципрокное торможение (рис. 3.10, В), т. е. процесс, основанный на том, что сигналы по одним и тем же афферентным путям обеспечива¬ют возбуждение одной группы нейронов, а через вставочные тормоз¬ные клетки вызывают торможение другой группы нейронов. Этот тип торможения называют иначе сопряженным. Проявляется реципрокное торможение, например, на уровне мотонейронов спинного мозга, ин¬нервирующих мышцы-антагонисты (сгибатели-разгибатели) конечно¬стей. Передача возбуждения к мотонейронам мышц-сгибателей одно¬временно через вставочные тормозные нейроны тормозит мотонейро¬ны мышц-разгибателей, благодаря чему только и возможно осуществ-ление сгибательного рефлекса; реципрокное торможение (рис. 3.10, В), т. е. процесс, основанный на том, что сигналы по одним и тем же афферентным путям обеспечива¬ют возбуждение одной группы нейронов, а через вставочные тормоз¬ные клетки вызывают торможение другой группы нейронов. Этот тип торможения называют иначе сопряженным. Проявляется реципрокное торможение, например, на уровне мотонейронов спинного мозга, ин¬нервирующих мышцы-антагонисты (сгибатели-разгибатели) конечно¬стей. Передача возбуждения к мотонейронам мышц-сгибателей одно¬временно через вставочные тормозные нейроны тормозит мотонейро¬ны мышц-разгибателей, благодаря чему только и возможно осуществ-ление сгибательного рефлекса;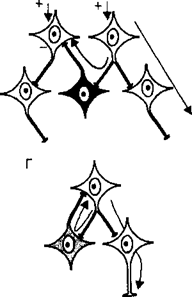 • возвратное торможение (рис. 3.10, Г), т. е. процесс торможения эф¬ферентных нейронов сигналами, возвращающимися к ним по колла- тералям аксонов через вставочные тормозные клетки. Возбуждение от одних нейронов поступает через ответвления на аксонах к вставоч¬ным тормозным нейронам, а аксоны этих клеток в свою очередь об¬разуют тормозные синапсы на телах первых нейронов. Примером яв¬ляется торможение Реншоу в спинном мозге, где импульсы от двига¬тельных нейронов посылаются по аксонам не только к скелетной мышце, но поступают через аксонные коллатерали к тормозным ин¬тернейронам Реншоу, а от них назад к двигательным нейронам, при¬водя к их торможению. Таким образом осуществляется короткая от¬рицательная обратная связь, не позволяющая возникнуть избыточно¬му возбуждению мотонейронов спинного мозга; • возвратное торможение (рис. 3.10, Г), т. е. процесс торможения эф¬ферентных нейронов сигналами, возвращающимися к ним по колла- тералям аксонов через вставочные тормозные клетки. Возбуждение от одних нейронов поступает через ответвления на аксонах к вставоч¬ным тормозным нейронам, а аксоны этих клеток в свою очередь об¬разуют тормозные синапсы на телах первых нейронов. Примером яв¬ляется торможение Реншоу в спинном мозге, где импульсы от двига¬тельных нейронов посылаются по аксонам не только к скелетной мышце, но поступают через аксонные коллатерали к тормозным ин¬тернейронам Реншоу, а от них назад к двигательным нейронам, при¬водя к их торможению. Таким образом осуществляется короткая от¬рицательная обратная связь, не позволяющая возникнуть избыточно¬му возбуждению мотонейронов спинного мозга;• 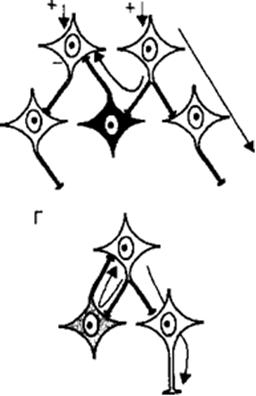 латеральное торможение (рис. 3.10, Б), т. е. процесс торможения группы нейронов, расположенной рядом с группой возбужденных клеток. Этот вид торможения распространен в сенсорных системах, где ответвления аксонов возбужденных нейронов образуют синапсы на вставочных тормозных нейронах, а аксоны последних оканчивают¬ся тормозными синапсами на соседних, рядом расположенных клет¬ках, получающих информацию от другого афферентного входа. Бла¬годаря явлению латерального торможения удается различать контраст в освещенности объектов, звуковые тоны и другие различия воспри¬нимаемых раздражителей; латеральное торможение (рис. 3.10, Б), т. е. процесс торможения группы нейронов, расположенной рядом с группой возбужденных клеток. Этот вид торможения распространен в сенсорных системах, где ответвления аксонов возбужденных нейронов образуют синапсы на вставочных тормозных нейронах, а аксоны последних оканчивают¬ся тормозными синапсами на соседних, рядом расположенных клет¬ках, получающих информацию от другого афферентного входа. Бла¬годаря явлению латерального торможения удается различать контраст в освещенности объектов, звуковые тоны и другие различия воспри¬нимаемых раздражителей;25. Вторичное торможение. Механизмы вторичного торможения. Не требует специальных тормозных структур, возникает в результате изменения функциональной активности обычных возбудимых структур, всегда связано с процессом возбуждения. Виды вторичного торможения: а) запредельное, возникающее при большом потоке информации, поступающей в клетку. Поток информации лежит за пределами работоспособности нейрона; б) пессимальное, возникающее при высокой частоте раздражения, в результате сильной деполяризации постсинаптической мембраны; в) парабиотическое, возникающее при сильно и длительно действующем раздражении; Парабиоз - явление пограничного состояния между гибелью и жизнью ткани (резко падают все свойства ткани, основное свойство - фазное изменение лабильности). Если парабиотический фактор продолжает действовать, ткань гибнет. г) торможение вслед за возбуждением, возникающее вследствие снижения функционального состояния нейронов после возбуждения. В конце акта возбуждения нейрона в нем может развиваться сильная следовая гиперполяризация. В то же время возбуждающий постсинаптический потенциал не может довести деполяризацию мембраны до критического уровня деполяризации, потенциалзависимые натриевые каналы не открываются и потенциал действия не возникает. д) торможение по принципу отрицательной индукции; е) торможение условных рефлексов 26. Принципы координационной деятельности ЦНС (принцип реципрокности, субординации, обратной афферентации, общего конечного пути, доминанты) При реализации сложных рефлекторных реакций, обеспечивающих регуляцию жизнедеятельности и приспособление организма к меняющимся условиям среды, необходимым условием является координация рефлексов, в основе которой лежит согласованное формирование и взаимодействие процессов возбуждения и торможения в центральной нервной системе. Основные принципы: • Принцип сопряженного торможения, или реципрокности, базируется на описанном выше одноименном виде торможения и означает возник¬новение при возбуждении одного центра одновременного торможе¬ния другого центра для обеспечения цепных рефлексов и сложной функции. Примером реципрокности может быть регуляция спинным мозгом противоположных по функциональному назначению мышц конечностей (рис. 3.11). Так, при возбуждении мотонейронов, иннер¬вирующих мышцы-сгибатели правой ноги, реципрокно тормозятся мотонейроны мышц-разгибателей этой ноги и возбуждаются мото¬нейроны мышц-разгибателей левой ноги. Формирующийся цепной характер рефлексов вслед за этим вызывает возбуждение мотонейро¬нов разгибателей правой ноги и реципрокно — торможение мотоней¬ронов сгибателей правой ноги и возбуждение мотонейронов сгибате¬лей левой ноги. Таким образом реципрокные взаимоотношения меж¬ду указанными рефлексами обеспечивают цепной шагательный реф¬лекс. • Принцип доминанты основан на открытом А. А. Ухтомским свойстве нервных центров. Доминантой называют общий принцип деятельно¬сти нервной системы, проявляющийся в виде господствующей в тече¬ние определенного времени системы рефлексов, реализуемых доми¬нирующими центрами, которые подчиняют себе или подавляют дея¬тельность других нервных центров и рефлексов. Нейроны доминини- рующих центров становятся более возбудимыми и способны эффек¬тивнее осуществлять пространственную и временную суммацию нервных импульсов. Синаптическое проведение к этим нейронам об¬легчено, и поэтому они могут возбуждаться и за счет «посторонних» импульсов от не имеющих прямых связей с доминирующими центра¬ми информационных каналов. Вследствие суммации многочисленных сигналов возбуждение нейронов, как и число возбужденных клеток в доминирующем центре, нарастает и осуществляемые им рефлектор¬ные реакции легко реализуются. Преобладание рефлексов домини¬рующего центра над другими рефлекторными актами становится осо¬бенно выраженным, поскольку через систему вставочных нейронов доминирующий центр сопряженно тормозит другие центры и теку¬щие рефлексы. Принцип доминанты позволяет концентрировать вни¬мание и строить поведение для достижения определенной намечен¬ной цели. • Принцип общего конечного пути, описанный Ч. Шеррингтоном, рас¬сматривался при изложении процесса конвергенции возбуждения. • Принцип субординации нервных центров (принцип подчинения) Прояв¬ляется в виде регулирующего влияния выше расположенных нервных центров на ниже расположенные. Так, двигательные центры головно¬го мозга управляют спинальными мотонейронами. Примером такого влияния является феномен центрального торможения спинальных рефлексов, открытый И. М. Сеченовым и получивший название се¬ченовского торможения. В эксперименте И. М. Сеченова раздраже¬ние зрительных бугров лягушки с помощью кристаллика поваренной соли (т. е. раздражение ретикулярной формации среднего мозга) при¬водило к торможению спинальных двигательных рефлексов, вызывае¬мых погружением лапки лягушки в слабый раствор кислоты. Следо¬вательно, торможение центров спинного мозга явилось следствием возбуждения центров среднего мозга. Прекращение этого тормозного контроля при перерыве цереброспинальных проводящих путей вызы¬вает резкое повышение возбудимости спинальных центров и гипер¬рефлексию. • Принцип обратной афферентации заключается в рецепторном воспри¬ятии результатов рефлекторного акта и проведении информации об¬ратно в структуры нервного центра, где она обрабатывается и сравни¬вается с сохраняющимися параметрами возбуждения. Обратная аффе- рентация реализуется в виде механизмов положительной или отрица¬тельной обратной связи. Таким образом, с помощью обратной аффе¬рентации нервные центры осуществляют непрерывный контроль эф¬фективности, целесообразности и оптимальности рефлекторной дея-тельности. 27. Вегетативная нервная система. Особенности вегетативной рефлекторной дуги. Вегетативный тонус. Вегетативная нервная система — отдел нервной системы, регулирующий деятельность внутренних органов, желёз внутренней и внешней секреции, кровеносных и лимфатических сосудов. Играет ведущую роль в поддержании постоянства внутренней среды организма и в приспособительных реакциях всех позвоночных. Анатомически и функционально вегетативная нервная система подразделяется на симпатическую и парасимпатическую. В симпатическом и парасимпатическом отделах имеются центральная и периферическая части. Центральную часть образуют тела нейронов, лежащих в спинном и головном мозге. Эти скопления нервных клеток получили название вегетативных ядер. Отходящие от ядер волокна, вегетативные ганглии, лежащие за пределами центральной нервной системы, и нервные сплетения в стенках внутренних органов образуют периферическую часть вегетативной нервной системы. Симпатические ядра расположены в спинном мозге. Отходящие от него нервные волокна заканчиваются за пределами спинного мозга в симпатических узлах, от которых берут начало нервные волокна. Эти волокна подходят ко всем органам. Парасимпатические ядра лежат в среднем и продолговатом мозге и в крестцовой части спинного мозга. Нервные волокна от ядер продолговатого мозга входят в состав блуждающих нервов. От ядер крестцовой части нервные волокна идут к кишечнику, органам выделения. Деятельность вегетативной нервной системы не зависит от воли человека. Рефлекторная дуга. Строение рефлекторных дуг вегетативного отдела отличается от строения рефлекторных дуг соматической части нервной системы. В рефлекторной дуге вегетативной части нервной системы эфферентное звено состоит не из одного нейрона, а из двух, один из которых находится вне ЦНС. В целом простая вегетативная рефлекторная дуга представлена тремя нейронами. Первое звено рефлекторной дуги — это чувствительный нейрон, тело которого располагается в спинномозговых узлах и в чувствительных узлах черепных нервов. Периферический отросток такого нейрона, имеющий чувствительное окончание — рецептор, берёт начало в органах и тканях. Центральный отросток в составе задних корешков спинномозговых нервов или чувствительных корешков черепных нервов направляется к соответствующим ядрам в спинной или головной мозг. Второе звено рефлекторной дуги является эфферентным, поскольку несёт импульсы из спинного или головного мозга к рабочему органу. Этот эфферентный путь вегетативной рефлекторной дуги представлен двумя нейронами. Первый из этих нейронов, второй по счёту в простой вегетативной рефлекторной дуге, располагается в вегетативных ядрах ЦНС. Его можно называть вставочным, так как он находится между чувствительным (афферентным) звеном рефлекторной дуги и вторым (эфферентным) нейроном эфферентного пути. Эффекторный нейрон представляет собой третий нейрон вегетативной рефлекторной дуги. Тела эффекторных (третьих) нейронов лежат в периферических узлах вегетативной нервной системы (симпатический ствол, вегетативные узлы черепных нервов, узлы внеорганных и внутриорганных вегетативных сплетений). Отростки этих нейронов направляются к органам и тканям в составе органных вегетативных или смешанных нервов. Заканчиваются постганглионарные нервные волокна на гладких мышцах, железа́х и в других тканях соответствующими концевыми нервными аппаратами. Вегетативный тонус. В естественных условиях симпатические и парасимпатические центры находятся в состоянии непрерывного возбуждения, получившего название «тонус». Характерной особенностью тонического влияния является длительное поддержание внешнего эффекта, который наиболее выраженно отражается на функциональном состоянии сосудистой стенки, сердечной мышцы, висцеральных органов в целом. Считают, что тонус ядерных образований формируется преимущественно благодаря притоку сенсорной информации из рефлексогенных зон, отдельных групп интероцепторов, а также соматических рецепторов. При этом не исключается и существование собственных водителей ритма — пейсмекеров, локализованных в основном в продолговатом мозге. Тоническое состояние можно непосредственно зарегистрировать в отдельных волокнах и клетках и оценить по показателям их активности. Тонус можно оценить еще и косвенно. В этом случае основным показателем является изменение деятельности органа после перерезки или электрической стимуляции иннервирующих его волокон (перерезка блуждающих нервов вызывает отчетливое учащение сердечного ритма). В целом тонус автономной нервной системы рассматривается как одно из проявлений гомеостатического состояния и одновременно один из механизмов его стабилизации. Преобладание в организме тонуса парасимпатической части автономной нервной системы именуется ваготонией, симпатической — симпатикотонией. 28. Влияние симпатического отдела ВНС на деятельность органов. Типы адренорецепторов и их функции. Влияние: На сердце — повышает частоту и силу сокращений сердца. На артерии — не влияет в большинстве органов, вызывает расширение артерий половых органов и мозга, сужение коронарных артерий и артерий лёгких. На кишечник — угнетает перистальтику кишечника и выработку пищеварительных ферментов. На слюнные железы — угнетает слюноотделение. На мочевой пузырь — расслабляет мочевой пузырь. На бронхи и дыхание — расширяет бронхи и бронхиолы, усиливает вентиляцию лёгких. На зрачок — расширяет зрачки. Типы адренорецепторов. α1- и β1-рецепторы локализуются в основном на постсинаптических мембранах и реагируют на действие норадреналина, выделяющегося из нервных окончаний постганглионарных нейронов симпатического отдела. α2- и β2-рецепторы являются внесинаптическими, а также имеются на пресинаптической мембране тех же нейронов. На α2-рецепторы действуют как адреналин, так и норадреналин. β2-рецепторы чувствительны в основном к адреналину. На α2-рецепторы пресинаптической мембраны норадреналин действует по принципу отрицательной обратной связи — ингибирует собственное выделение. При действии адреналина на β2-адренорецепторы пресинаптической мембраны выделение норадреналина усиливается. Кратко охарактеризовать значение рецепторов можно следующим образом: α1 — локализуются в артериолах, стимуляция приводит к спазму артериол, повышению давления, снижению сосудистой проницаемости и уменьшению эксудативного воспаления. α2 — главным образом пресинаптические рецепторы, являются «петлёй обратной отрицательной связи» для адренэргической системы, их стимуляция ведёт к снижению артериального давления. β1 — локализуются в сердце, стимуляция приводит к увеличению частоты (положительный хронотропный эффект) и силы сердечных сокращений,(положительный инотропный эффект) кроме того, приводит к повышению потребности миокарда в кислороде и повышению артериального давления. Также локализуются в почках, являясь рецепторами юкстагломерулярного аппарата. β2 — локализуются в бронхиолах, стимуляция вызывает расширение бронхиол и снятие бронхоспазма. Эти же рецепторы находятся на клетках печени, воздействие на них гормона вызывает гликогенолиз и выход глюкозы в кровь. β3 — находятся в жировой ткани. Стимуляция этих рецепторов усиливает липолиз и приводит к выделению энергии, а также к повышению теплопродукции. 29. Влияние парасимпатического отдела ВНС на деятельность органов. Типы холинорецепторов и их функции. Влияние парасимпатического отдела: На сердце — уменьшает частоту и силу сокращений сердца. На артерии — расширяет артерии. На кишечник — усиливает перистальтику кишечника и стимулирует выработку пищеварительных ферментов. На слюнные железы — стимулирует слюноотделение. На мочевой пузырь — сокращает мочевой пузырь. На бронхи и дыхание — сужает бронхи и бронхиолы, уменьшает вентиляцию лёгких. На зрачок — сужает зрачки. На постсинаптической мембране ацетилхолин связывается с М-холино- рецепторами нескольких типов, что и определяет различия в характере ре¬акций органов на этот медиатор. По чувствительности к различным фар¬макологическим препаратам выделяют М1 – М4 холинорецепторы, которые локализуются: M1-холинорецепторы — в вегетативных ганглиях и ЦНС; М2-холинорецепторы — в сердце, в гладких мышцах желудочно-кишеч¬ного тракта; М3- холинорецепторы — в гладких мышцах, в большинстве экзокринных желез. М4-холинорецепторы изучены мало. На постсинаптической мембране ацетилхолин образует с М-холиноре- цепторами медиатор-рецепторный комплекс, который активирует натрие¬вые каналы и возбуждает гладкомышечные и секреторные клетки желудоч¬но-кишечного тракта, клетки гладких мышц бронхов, мочевого пузыря и мочеточника. Возбуждение М-холинорецепторов вызывает: сужение бронхов, увеличе¬ние слюноотделения и слезоотделения, увеличение моторики и секреции соков в желудочно-кишечном тракте, а также сужение зрачка. Этому спо¬собствует активация вторичных посредников — инозитол-3-фосфата и ио¬низированного кальция. В клетках проводящей системы сердца и гладких мышц сосудов поло¬вых органов ацетилхолин активирует калиевые каналы и выходящий ток калия, что приводит к гиперполяризации постсинаптических мембран. В результате этого происходят торможение ритма сердца, снижение проводи¬мости и возбудимости в миокарде, расширение артерий половых органов. Одновременно в клетках активируется система вторичных посредников — цГМФ. М-холинорецепторы блокирует атропин, который тормозит пара¬симпатическую стимуляцию сокращения гладких мышц и парасимпатиче¬ское торможение деятельности сердца. |