ответы на физиологию. 1. Современные представления о строении и функции мембран
 Скачать 0.69 Mb. Скачать 0.69 Mb.
|

|
7. Изменение возбудимости во время генерации потенциала действия. Понятие о рефрактерности, причины её возникновения. В фазу деполяризации потенциала действия потенциалзависимые натриевые ионные каналы кратковременно открываются, но затем инактивируются h-ворота (см. рис. 2.9). В период инактивации натриевых ионных каналов возбудимые клетки не способны реагировать повышением натриевой проницаемости на повторный стимул. Поэтому во время фазы деполяризации мембрана не может генерировать потенциал действия в ответ на действие пороговых или сверхпороговых раздражителей. Это состояние называется абсолютной рефрактерностъю> время которой составляет в нервных волокнах 0,5—1,0 мс, а в скелетных мышечных клетках —в среднем 2 мс. Период абсолютной рефрактерности заканчивается после того, как снижается количество инактивированных натриевых каналов и постепенно увеличивается количество натриевых каналов, находящихся в закрытом состоянии. Происходят эти процессы во время фазы реполяризации, когда уменьшению количества потенциалзависимых натриевых ионных каналов, находящихся в состоянии инактивации, соответствует период относительной рефрактерности. Период относительной рефрактерности характеризуется тем, что лишь некоторая часть потенциалзависимых натриевых ионных каналов переходит в закрытое состояние, а в силу этого порог возбудимости мембраны клетки имеет более высокие значения, чем в исходном состоянии. Поэтому возбудимые клетки в период относительной рефрактерности могут генерировать потенциалы действия, но при воздействии на них раздражителей сверхпороговой силы. Однако из-за небольшого количества потенциалзависимых натриевых ионных каналов, находящихся в закрытом состоянии, амплитуда генерируемых при этом потенциалов действия будет меньше, чем в условиях исходной возбудимости нервной или мышечной клетки. В клетках возбудимых тканей максимальное количество генерируемых потенциалов действия в единицу времени обусловлено двумя факторами: длительностью потенциала действия и длительностью периода абсолютной рефрактерности после каждого импульса. На этом основании в физиологии формулируется современное понятие лабильности: чем меньше период абсолютной рефрактерности при возбуждении возбудимой ткани, тем выше ее функциональная подвижность или лабильность, тем больше в ней генерируется потенциалов действия в единицу времени. При непрерывной стимуляции нерва электрическим током лабильность нерва зависит от частоты и силы раздражения. В зависимости от частоты и силы раздражения нерва сокращение иннервируемой им мышцы может быть максимальной или минимальной амплитуды. Эти явления были названы соответственно оптимумом и пессимумом (Н. Е. Введенский). Максимальное (оптимально большое) сокращение мышцы возникает в том случае, если каждый последующий электрический стимул действует на нерв в периоде его состояния супернормальной возбудимости после предыдущего потенциала действия. Минимальное (или пессимальное) сокращение мышцы возникает в том случае, если каждый последующий электрический стимул действует на нерв, находящийся в периоде относительной рефрактерности после предыдущего потенциала действия. Поэтому значения оптимальной частоты раздражения нерва всегда меньше, чем значения пессимальной частоты раздражения. 8. Общие свойства возбудимых тканей. Возбудимость, критерии её оценки. Свойства: 1) Возбудимость – это способность отвечать возбуждением (генерацией потенциала действия) в ответ на действие раздражителей любой природы. 2) проводимость – способность ткани передавать возникшее возбуждение за счет электрического сигнала от места раздражения по длине возбудимой ткани; 3) Рефрактерность – временное снижение возбудимости одновременно с возникшим в ткани возбуждением. Рефрактерность бывает абсолютной (нет ответа ни на какой раздражитель) и относительной (возбудимость восстанавливается, и ткань отвечает на подпороговый или сверхпороговый раздражитель); 4) лабильность (функциональная подвижность) - способность ткани воспроизводить определенное число волн возбуждения в единицу времени в точном соответствии с ритмом наносимых раздражений. Это свойство характеризует скорость возникновения возбуждения. Показатель лабильности: максимальное количество волн возбуждения в данной ткани: нервные волокна - 500-1000 импульсов в секунду, мышечная ткань - 200-250 импульсов в секунду, синапс - 100-125 импульсов в секунду. Лабильность зависит от уровня обменных процессов в ткани, возбудимости, рефрактерности. 5) Сократимость (для мышечных волокон!) - способность мышечных волокон укорачиваться или изменять свое напряжение. - Растяжимость - ткань можно растянуть, не повредив ее - Эластичность - способность принимать исходную форму после растяжения. Критерии оценки возбудимости: - Пороговая сила – это наименьшая сила раздражителя, способная вызвать ПД при неограниченном во времени действии раздражителя. При использовании в качестве раздражителя электрический ток, его пороговая сила равна 1 реобазе. Если возбудимость ткани высока, пороговая сила раздражителя мала. - Пороговое время – это минимальное время, в течение которого должен действовать на ткань раздражитель пороговой силы, чтобы вызвать ее возбуждение. Хронаксия – наименьшее время, в течение которого должен действовать ток в две реобазы, чтобы вызвать возбуждение. - критический уровень деполяризации - величина мембранного потенциала, при достижении которой возникает потенциал действия. Хронаксия как мера возбудимости. Поскольку возникновение возбужде¬ния в нервной или мышечной ткани обусловлено силой и продолжитель¬ностью раздражения, то величина полезного времени не является точной мерой функционального состояния возбудимых тканей. Это обусловлено следующими обстоятельствами. Взаимодействие между силой и длительно¬стью стимула, вызывающего возбуждение нервной или мышечной ткани характеризуется кривой «сила-длительность» (рис. 2.6). Для определения величины полезного времени возбудимых тканей необходимо построение подобной кривой, что требует многочисленных воздействий электриче¬ским током на возбудимые ткани и проведения многократных измерений этого показателя. Однако при минимальной силе раздражения, способного вызвать возбуждение в нервной или мышечной ткани, значение полезного времени соответствует, как показано на рисунке, точке Р. Вправо от точки Р кривая становится параллельной оси абсцисс, а следовательно, возбуж-дение в нервной или мышечной ткани может возникать при любой дли¬тельности действующего раздражителя (закон раздражения) и становится невозможным по полезному времени точно определить функциональное состояние возбудимых тканей. Поэтому французским ученым Лапиком экспериментально был выработан способ получения более точной характе¬ристики возбудимости нервной или мышечной ткани как функции време¬ни. Согласно этому предложению, минимальная сила электрического раз¬дражения (названа реобазой), которая способная вызвать возбуждение тка¬ни независимо от длительности действия электрического тока, должна быть удвоена, чтобы установить полезное время. Минимальное время, в течение которого ток силы, равной двум реобазам, должен действовать на возбудимую ткань, чтобы вызвать минимальный эффект возбуждения, бы¬ло названо хронаксией. Хронаксия (обозначается буквой о) является мерой возбудимости нервной или мышечной ткани и отражает скорость возник¬новения возбуждения в этих тканях. Значение хронаксии, как показано на кривой рисунка, находится в области больших значений длительности раз¬дражения (восходящая часть кривой). Так, у мышц человека хронаксия со¬ставляет 0,1—0,7 мс. При этом хронаксия мышц-сгибателей (0,08—0,16 мс) равна вдвое меньше, чем хронаксия мышц-рагибателей (0,16—0,32 мс). Указанные величины продолжительности хронаксии мышц-сгибателей и разгибателей у человека обусловлены возбудимостью нервных волокон, иннервирующих эти мышцы. От возбудимости нервной или мышечной ткани, как это будет изложено в разделах настоящей главы, зависит, на¬пример, скрытый период появления возбуждения в возбудимых тканях, длительность и интенсивность возникающего в них возбуждения. Поэтому, в наиболее общем виде, чем меньше хронаксия, тем сильнее в ткани про¬цесс возбуждения, тем меньше скрытый период возникновения возбужде¬ния. Определение хронаксии производится в клинической практике для оценки функционального состояния мышц при их повреждении. При этом чем больше хронаксия, тем хуже функциональное состояние поврежден¬ных мышц человека. 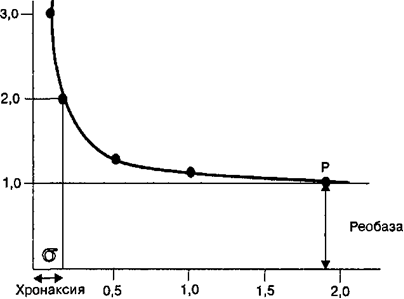 Мембрана клеток возбудимых тканей является высокочувствительной к действию электриче-ского тока. Для того, чтобы электрический ток вызвал возбуждение клетки, его сила должна быть достаточной силы. Сила тока, которая вызывает возбуждение нервной и мышечной тка¬ни, называется реобазой. Время, необходимое для того, чтобы ток силой удвоенной реобазы, вызвал возбуждение в ткани, называется хронаксией. Кривая сила—длительность отражает тот факт, что для достижение порога возбуждения в возбудимых тканях необходима пороговая скорость нарастания силы раздражения. 9. Проведение нервного импульса по безмиелиновым и миелиновым нервным волокнам. Законы проведения возбуждения. Немиелинизированные волокна Проведение потенциала действия по немиелинированному нервному во¬локну происходит путем активации потенциалзависимых натриевых ион¬ных каналов участка мембраны волокна, прилегающего к тому месту, где возник потенциал действия. При этом между возбужденным и невозбуж¬денным участками мембраны нервного волокна возникают локальные электрические токи, которые вызывают деполяризацию мембраны невоз¬бужденного участка до критического уровня, и только после этого в мембране нервного волокна мгновенно открываются потенциалзависимые на¬триевые каналы. Потенциал действия, таким образом, генерируется в по¬следующем участке нервного волокна. 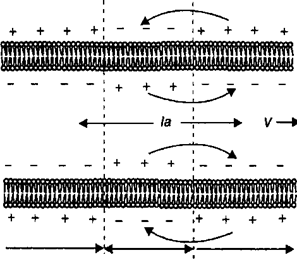 Распространение потенциа¬ла действия по немиелинизированно- му нервному волокну. При деполяризации мембраны нервного волокна возникают продольные токи, которые распространяются по аксоплазме и направлены от активного участка генерации потенциала действия в обе стороны нервного волокна. Продольные токи (1а) вызывают открытие натриевых потенциалзависимых ионных каналов в участке мембраны, который имеет нормальную возбудимость и вызывают в этом месте генерацию потенциала действия. Рефрактерный участок невозбудим, поскольку натриевые ионные каналы находятся в инактивированом состоянии. В результате потенциал действия генерируется на соседнем участке мембраны и движется по нервному волокну в одном направлении. Распространение потенциа¬ла действия по немиелинизированно- му нервному волокну. При деполяризации мембраны нервного волокна возникают продольные токи, которые распространяются по аксоплазме и направлены от активного участка генерации потенциала действия в обе стороны нервного волокна. Продольные токи (1а) вызывают открытие натриевых потенциалзависимых ионных каналов в участке мембраны, который имеет нормальную возбудимость и вызывают в этом месте генерацию потенциала действия. Рефрактерный участок невозбудим, поскольку натриевые ионные каналы находятся в инактивированом состоянии. В результате потенциал действия генерируется на соседнем участке мембраны и движется по нервному волокну в одном направлении.Рефракт.уч. Акт.уч. Норм.возб. Миелинизированные волокна В миелинизированном нервном волокне потенциалы действия генериру¬ются только в области перехвата Ранвье, т. е. той части мембраны, которая не покрыта шванновскими клетками. С одной стороны, это обусловлено тем, что цитоплазма шванновской клетки содержит липид — сфингомиелин, который уменьшает поток ионов через мембрану нервного волокна при¬мерно в 5000 раз и снижает ее емкость в 50 раз. С другой стороны, в об¬ласти перехвата Ранвье в мембране нервного волокна имеется наибольшее число потенциалзависимых натриевых ионных каналов, а сама мембрана обладает нормальным уровнем возбудимости. При нанесении точечного раздражения на миелинизированное нервное волокно потенциал действия генерируется в зоне перехвата Ранвье и возникают электрические токи, ко¬торые текут вдоль силовых линий от плюса к минусу потенциала на мем¬бране (рис. 2.14). Одновременно в аксоплазме нервного волокна возника¬ют продольные токи, которые направлены от места генерации потенциала действия в обе стороны (1а). Продольные токи вызывают открывание по-тенциалзависимых натриевых ионных каналов, а следовательно, движение ионов Na+ через мембрану и генерацию потенциала действия в соседних перехватах Ранвье, минуя часть нервного волокна, покрытого шванновски¬ми клетками. Поскольку потенциалы действия передаются (перепрыгива¬ют) от одного перехвата Ранвье к другому, то механизм проведения потен¬циалов действия в миелинизированных нервных волокнах получил название сальтаторного. В миелинизированных нервных волокнах реполяриза¬ция мембраны после потенциала действия происходит с очень высокой скоростью. В результате нервные волокна имеют высокую функциональ¬ную лабильность и способны проводить значительное число потенциалов действия в единицу времени. Скорость распространения потенциалов дей¬ствия в зависимости от диаметра и типа миелинизированных нервных во¬локон чрезвычайно высока и варьирует от 6 до 120 м/с. В толстых миели¬низированных нервных волокнах скорость проведения возбуждения про¬порциональна диаметру волокна, а проводимость нервного волокна имеет обратную зависимость от его диаметра. Законы проведения возбуждения по нервному волокну Проведение возбуждения по нервному волокну возможно при условии его структурной целостности и физиологической непрерывности (закон физио-логической непрерывности). Физиологическая непрерывность нерва может быть нарушена, например, при сдавлении нерва без его структурного по¬вреждения, что препятствует проведению потенциалов действия. При про¬ведении возбуждения по нервному волокну потенциал действия не распро¬страняется с одного волокна на другое, например рядом расположенное (закон изолированного проведения). Несмотря на то что нервные волокна проводят потенциалы действия либо в направлении тела нейрона (центро¬стремительно), либо от тела нервной клетки к окончаниям аксона (цен- тробежно), отдельное нервное волокно обладает двусторонней проводимо¬стью (закон двустороннего проведения). При искусственном электрическом раздражении в любой точке по ходу нервного волокна может возникать потенциал действия и распространяться как центростремительно, так и центробежно. 10. Характеристика волокон А, В, С.
В зависимости от диаметра, наличия или отсутствия миелиновой обо¬лочки и скорости проведения нервных импульсов нервные волокна делят на три класса: А (четырех типов), В и С (табл. 3.1). Миелинизированные волокна более толстые, скорость проведения сигналов по ним существенно больше. Так, волокна A-типов обеспечивают проведение сигналов при рефлекторной регуляции скелетных мышц. Немиелинизированные тонкие волокна С-типа участвуют в проведении сигналов, регулирующих деятель¬ность внутренних органов, скорость проведения сигнала по ним — самая низкая. 11. Синапс. Строение, классификация синапсов. Синапсом называется место контакта нервной клетки с другим нейроном или исполнительным органом. Все синапсы делятся на следующие группы: 1. По механизму передачи: а. электрические. В них возбуждение передается посредством электрического поля. Поэтому оно может передаваться в обе стороны. Их в ЦНС мало; б. химические. Возбуждение через них передается с помощью ФАВ – нейромедиатора. Их в ЦНС большинство; в. смешанные (электрохимические). 2. По локализации: а. центральные, расположенные в ЦНС; б. периферические, находящиеся вне ее. Это нервно-мышечные синапсы и синапсы периферических отделов вегетативной нервной системы. 3. По физиологическому значению: а. возбуждающие; б. тормозные. 4. В зависимости от нейромедиатора, используемого для передачи: а. холинергические – медиатор ацетилхолин (АХ); б. адренергические – норадреналин (НА); в. серотонинергические – серотонин (СТ); г. глицинергические – аминокислота глицин (ГЛИ); д. ГАМК-ергические – гамма-аминомасляная кислота (ГАМК); е. дофаминергические – дофамин (ДА); ж. пептидергические – медиаторами являются нейропептиды. В частности роль нейромедиаторов выполняют вещество Р, опиоидный пептид в-эндорфин и др. Предполагают, что имеются синапсы, где функции медиатора выполняют гистамин, АТФ, глутамат, аспартат, ряд местных пептидных гормонов. 5. По месту расположения синапса: а. аксо-дендритные (между аксоном одного и дендритом второго нейрона); б. аксо-аксональные; в. аксо-соматические; г. дендро-соматические; д. дендро-дендритные. Наиболее часто встречаются три первых типа. Строение всех химических синапсов имеет принципиальное сходство. Например, аксо-дендритный синапс состоит из следующих элементов: 1. пресинаптическое окончание или терминаль (конец аксона); 2. синаптическая бляшка, утолщение окончания; 3. пресинаптическая мембрана, покрывающая пресинаптическое окончание; 4. синаптические пузырьки в бляшке, которые содержат нейромедиатор; 5. постсинаптическая мембрана, покрывающая участок дендрита, прилегающий к бляшке; 6. синаптическая щель, разделяющая пре- и постсинаптическую мембраны, шириной 10-50 нМ; 7. хеморецепторы – белки, встроенные в постсинаптическую мембрану и специфичные для нейромедиатора. Например, в холинергических синапсах это холинорецепторы, адренергических – адренорецепторы и т.д. Простые нейромедиаторы синтезируются в пресинаптических окончаниях, пептидные – в соме нейронов, а затем по аксонам транспортируются в окончания. |