ответы на физиологию. 1. Современные представления о строении и функции мембран
 Скачать 0.69 Mb. Скачать 0.69 Mb.
|

|
12. Механизм передачи возбуждения в синапсах (электрических, химических). Передача возбуждения в электрическим синапсе осуществля¬ется аналогично его проведению по нервным волокнам с помощью мест¬ных токов, возникающих между деполяризованным участком мембраны нервного волокна и поляризованным участком мембраны нервной клетки. Локальные токи деполяризуют мембрану нейрона до критического уров¬ня, после чего возникает спонтанный процесс регенеративной деполяри¬зации. Электрическая передача возбуждения осуществляется с высокой скоростью, близкой к скорости проведения возбуждения по нервным во¬локнам. В химическом синапсе импульс предается с помощью пре- и постсинаптического меха¬низмов. Пресинаптический механизм проведения возбуждения Пресинаптический механизм проведения возбуждения осуществляет трансформацию энергии нервных импульсов в кванты выделяющегося ме¬диатора. В фазу деполяризации потенциала действия в мембране нервной терминали открываются потенциалзависимые кальциевые ионные каналы, и ионы кальция по концентрационному градиенту поступают из внекле¬точной среды внутрь нервного окончания. Ионы Са2+ вблизи внутренней поверхности пресинаптической мембраны образуют микродомен и связываются со специфическим везикулярным белком каль¬циевым сенсором — синаптотагмином. Последовательное взаимодействие специфических белков экзоцитоза, локализованых на мембране везикул (синапсин, синаптотагмин, синаптобревин) и на пресинаптической мем¬бране (синтаксин, синапсоассоциированный белок) связывает везикулу с активной зоной пресинаптической мембраны, формируя секретосому. При этом мембрана везикулы полностью сливается с пресинаптической мем¬браной, и медиатор освобождается в синаптическую щель через пору. В си¬напсах нервной системы экзоцитоз медиаторов может проходит через по¬стоянную или временную пору. В первом случае происходят полное слияние мембран везикулы и пресинаптического окончания и высвобождение всего количества молекул медиатора из везикул. Во втором случае не про¬исходит полного слияния мембран везикулы и пресинаптической термина- ли. При образовании временной поры освобождается только часть медиа¬тора в синаптическую щель, а затем везикула отделяется от активной зоны и возвращается в цитоплазму нервной терминали. Этот механизм позволя¬ет одной и той же везикуле без пополнения запаса медиатора неоднократ¬но участвовать в экзоцитозе. После высвобождения молекулы нейротрансмиттера диффундируют от мест экзоцитоза к постсинаптической мембране. Во время диффузии часть медиатора может подвергаться ферментному разрушению в синаптической щели или захватываться обратно внутрь нервного окончания. Постсинап¬тической мембраны могут достигать все или только часть молекул ней¬ротрансмиттеров, что обусловливает соответственно большее или меньшее количество передаваемых через аксосоматический синапс импульсов. Постсинаптические рецепторы делятся на две большие группы: 1) рецепторы, сопряженные с ионными каналами, или ионотропные рецепторы; 2) рецепторы, сопряженные с внутриклеточными ферментативными системами, или метаботропные рецепторы. Ионотропные рецепторы Это ионные каналы, на наружной поверхности которых имеются собственно рецепторы медиаторов (отсюда синонимы— хемочувствительные каналы; рецепторы-каналы; рецепторы, сопряженные с ионными каналами). Общие этапы передачи сигнала посредством таких рецепторов следующие. 1. Медиатор активирует рецептор. 2. Активация рецептора приводит к открыванию (реже — закрыванию) ионного канала. 3. Меняется поток через канал ионов, для которых данный канал избирательно проницаем. 4. Поток ионов вызывает изменение заряда мембраны; возникает постсинаптический потенциал, обладающий всеми свойствами местных потенциалов (распространяется с затуханием; подчиняется закону силовых отношений; не обладает рефрактерностью; способен к суммации). В зависимости от того, для каких ионов проницаемы связанные с рецепторами каналы, могут возникать два вида постсинаптических потенциалов (ВПСП И ТПСП). Метаботропные рецепторы — это мембранные молекулярные рецепторы, которые передают внешний химический управляющий сигнал внутрь клетки, запуская каскад биохимических реакций и воздействуют на клеточный метаболизм. Таким образом, метаботропные рецепторы влияют на состояние клетки опосредованно, а не прямо. Метаботропные рецепторы представляют собой молекулярную систему, состоящую, по крайней мере, из трех белков: 1) собственно рецепторного белка, связывающегося с нейромедиатором; 2) так называемого G-белка, передающего сигнал с рецепторного белка внутри клетки, и 3) белка-эффектора, который является ферментом, катализирующим образование внутриклеточного низкомолекулярного регулятора, так называемого вторичного мессенджера. Таким образом, задача метаботропного рецептора - запустить в клетке работу вторичных мессенджеров, которые изменят метаболизм клетки. Итак, при связывании со своим биолигандом (модулятором) метаботропные рецепторы активируют внутри клетки вещество-посредник (вторичный мессенджер), который запускает каскад ферментативных реакций. Первым таким посредником чаще всего бывает G-белок, связанный с метаботропным рецептором на внутренней стороне мембраны. Конечным результатом активации метаботропных рецепторов может быть опосредованное открытие ионных каналов клеточной мембраны, что, в свою очередь, позволяет ионам проникать через них в клетку или, наоборот, выходить через них в межклеточное пространство. 13. Медиаторы синаптической передачи, их виды и свойства. Нейромедиа́торы (нейротрансмиттеры, посредники) — биологически активные химические вещества, посредством которых осуществляется передача электрического импульса от нервной клетки через синаптическое пространство между нейронами, а также, например, от нейронов к мышечной ткани. Нервный импульс, поступающий в пресинаптическое окончание, вызывает освобождение в синаптическую щель медиатора. Молекулы медиаторов реагируют со специфическими рецепторными белками клеточной мембраны, инициируя цепь биохимических реакций, вызывающих изменение трансмембранного тока ионов, что приводит к деполяризации мембраны и возникновению потенциала действия. 1)Аминокислоты - ГАМК — важнейший тормозной нейромедиатор ЦНС. - Глицин — как нейромедиаторная аминокислота, проявляет двоякое действие. Глициновые рецепторы имеются во многих участках головного мозга и спинного мозга. Связываясь с рецепторами, глицин вызывает «тормозящее» воздействие на нейроны, уменьшают выделение из нейронов «возбуждающих» аминокислот, таких как глутамат, и повышают выделение ГАМК. Также глицин связывается со специфическими участками NMDA-рецепторов и, таким образом, способствует передаче сигнала от возбуждающих нейротрансмиттеров глутамата и аспартата. В спинном мозге глицин приводит к торможению мотонейронов, что позволяет использовать глицин в неврологической практике для устранения повышенного мышечного тонуса. - Глутаминовая кислота (глутамат) — наиболее распространенный возбуждающий нейротрансмиттер в нервной системе позвоночных, в нейронах мозжечка и спинного мозга - Аспарагиновая кислота (аспартат) — возбуждающий нейромедиатор в нейронах коры ГМ. 2) Катехоламины - Адреналин — относят к возбуждающим нейромедиаторам. - Норадреналин — считается одним из важнейших «медиаторов бодрствования». Норадренергические проекции участвуют в восходящей ретикулярной активирующей системе. Является медиатором как голубоватого пятна (лат. locus coeruleus) ствола мозга, так и окончаний симпатической нервной системы. Количество норадренергических нейронов в ЦНС невелико (несколько тысяч), но у них весьма широкое поле иннервации в головном мозге. - Дофамин — является одним из химических факторов внутреннего подкрепления и служит важной частью «системы поощрения» мозга, поскольку вызывает чувство удовольствия (или удовлетворения), чем влияет на процессы мотивации и обучения. 3) Другие моноамины - Серотонин — играет роль нейромедиатора в ЦНС. Серотонинергические нейроны группируются в стволе мозга: в варолиевом мосту и ядрах шва. От моста идут нисходящие проекции в спинной мозг, нейроны ядер шва дают восходящие проекции к мозжечку, лимбической системе, базальным ганглиям, коре. При этом нейроны дорсального и медиального ядер шва дают аксоны, различающиеся морфологически, электрофизиологически, мишенями иннервации и чувствительностью к некоторым нейротоксичным агентам, например, метамфетамину. Сон-бодрствование, настроение, аппетит, память, регуляция болевых импульсов, сексуальное поведение, регенерация мозга - Гистамин —содержатся в ЦНС 4) Другие представители - Ацетилхолин — осуществляет нервно-мышечную передачу, а также основной нейромедиатор в парасимпатической нервной системе. - Анандамид — является нейротрансмиттером и нейрорегулятором, который играет роль в механизмах происхождения боли, депрессии, аппетита, памяти, репродуктивной функции. - Таурин — играет роль нейромедиаторной аминокислоты, тормозящей синаптическую передачу, обладает противосудорожной активностью, оказывает также кардиотропное действие. 14. Механизмы формирования ВПСП И ТПСП Результатом взаимодействия медиаторов с ионотропными рецепторами постсинаптической мембраны является ее кратковременная деполяризация или гиперполяризация постсинаптической мембраны. Деполяризация постсинаптической мембраны в области одного синапса называется возбу¬ждающим постсинаптическим потенциалом (ВПСП). Химические синап¬сы, в которых при передаче возбуждения возникают ВПСП, называются возбуждающими. По аналогии медиатор, вызывающий возникновение ВПСП, называется возбуждающим медиатором. Возникновение ВПСП обусловлено открытием преимущественно натриевых ионных каналов. При ВПСП происходит деполяризация постсинаптической мембраны и уменьшение порога ее возбудимости. Амплитуда ВПСП прямо зависит от количества молекул медиатора взаимодействующих с рецепторами постси¬наптической мембраны, а также последовательностью выделения квантов медиатора из пресинаптического окончания. Результатом взаимодействия медиаторов с ионотропными рецепторами хемозависимых ионных каналов постсинаптической мембраны может быть ее кратковременная гиперполяризация. В результате понижается возбуди¬мость постсинаптического нейрона и в нем не происходит генерация по¬тенциала действия. Гиперполяризация постсинаптической мембраны в об¬ласти одного синапса называется тормозным постсинаптическим потен¬циалом (ТПСП), такие химические синапсы называются тормозными. По аналогии медиаторы, вызывающие ТПСП, называются тормозными медиа¬торами. Взаимодействие тормозного медиатора с рецепторами постсинап¬тической мембраны открывает хлорные и калиевые ионные каналы, что резко повышает ионную проводимость постсинаптической мембраны для ионов хлора (1а) или калия (/*). В соответствии с электрохимическими градиентами ионов калия и хлора по обе стороны от постсинаптической мембраны, ТПСП представляет собой сдвиг потенциала на постсинаптиче¬ской мембране в сторону увеличения его величины. В результате происхо¬дит гиперполяризация постсинаптической мембраны и увеличение порога ее возбудимости. При передаче возбуждения через аксосоматические синапсы на мембра¬не одного нейрона одновременно возникают ВПСП и ТПСП. При этом происходит сложение (суммация) амплитуды ВПСП и ТПСП многих си¬напсов. Если в условиях суммации ВПСП и ТПСП суммарная амплитуда ВПСП достигает критического уровня деполяризации, то в мембране ак¬сонного холмика нейрона электротонически открываются потенциалзави¬симые натриевые ионные каналы и генерируется потенциал действия. В результате возбуждение передается от нервных терминалей пресинаптиче¬ского нейрона на постсинаптический нейрон. 15. Молекулярные механизмы сокращения и расслабления скелетной мышцы. Электромеханическое сопряжение. 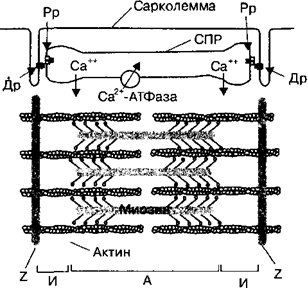 Во время укорочения мышцы уменьшается длина саркомеров. При этом филаменты актина и миозина, входящие в состав саркомеров, не изменяют своей длины. Уменьшение длины саркомеров происходит в результате движения (скольжения) тонких филаментов с двух сторон саркомера к его Во время укорочения мышцы уменьшается длина саркомеров. При этом филаменты актина и миозина, входящие в состав саркомеров, не изменяют своей длины. Уменьшение длины саркомеров происходит в результате движения (скольжения) тонких филаментов с двух сторон саркомера к его центру относительно толстых миозиновых филаментов. Цикл поперечного мостика во время сокращения мышцы - это связывание, укорочение, высвобождение и вновь связывание поперечного мостика миозина с актиновым филаментом. В состоянии покоя поперечный мостик расположен параллельно мио- зиновому стержню, а его головка — перпедикулярно относитель¬но последнего, и не прикреплена к актиновому филаменту (стадия 1). По¬сле того, как ионы Са2+ высвобождаются из саркоплазматического ретику¬лума (описано ниже) и активируют сокращение мышцы, активновый фи- ламент может связываться с миозиновой головкой (стадия 2). После свя¬зывания головка наклоняется (стадия 3), используя энергию, заключенную в миозин-АТФазном комплексе головки. Это является основной силой по¬перечного мостика, которая растягивает его эластический компонент для того, чтобы произвести энергию движения вдоль активного филамента (стадия 4). В результате головка мостика совершает движение на 40° к цен¬тру саркомера и перемещает при этом на 10 нм нить актина, В это время от головки высвобождаются АДФ и монофосфат, что необходимо для за-хвата головкой миозина новой молекулы АТФ. Только после этого головка миозина отделяется от молекулы актина, необходимой для размыкания по-перечного мостика с актином (положение 5). Цикл будет продолжаться до тех пор, пока концентрация ионов Са++ в межфибриллярном пространстве не снижается до подпороговой величины, что приведет к расслаблению мышцы, или пока не будут ис¬черпаны запасы АТФ. В последнем случае в мышце возникает спазм или мышечная контрактура (rigor), при которой в течение нескольких секунд или минут отсутствует расслабление мышцы. Таким образом, функция АТФ проявляется в двух точках процесса со¬кращения мышцы. Во-первых, АТФ является источником энергии для со¬кращения. Во-вторых, АТФ отсоединяет поперечной мостик для того, что¬бы повторялся цикл поперечных мостиков «замыкание—сокращение—раз¬мыкание». Расслабление скелетной мышцы Расслабление скелетной мышцы начинается только после того, как кон¬центрация ионов Са2+ в саркоплазме мышечных клеток снижается ниже порогового уровня, т. е. ниже 10 мкМ/л, или 10"5 М. Расслабление мышцы обусловливают три процесса: 1. Отсоединение (диссоциация) ионов Са2+ от субъединицы С тропонина. 2. Диффузия ионов Са2+ в сторону сарко¬плазматического ретикулума. 3. Активный транспорт ионов Са2+ внутрь СПР при участии Са2+-АТФазы. В начальный момент расслабления длина мышцы остается постоянной. Это происходит из-за того, что первоначально происходит расслабление саркомеров, находящихся на концах мышц, в то время как центральные саркомеры остаются укороченными. Это состояние характерно лишь для начального момента мышечного расслабления, а именно фазы изометрического расслабления. Процесс расслабления во всех саркомерах усиливается после того как сила сокращения мышцы уменьшается примерно на 20— 30 %. Это обусловлено тем, что во время расслабления мышцы уменьшает¬ся подвижность тропомиозина по поверхности белковых цепей актина примерно с той же скоростью, что и сила сокращения. В результате чем меньше сила сокращения мышцы, тем меньше открыто мест связывания на актиновых филаментах для поперечных мостиков миозина. Электромеханическое сопряжение - процесс, при котором потенциалы действия, генерируемые на сарколемме в области нервно-мышечного синапса, активируют сокращение мышцы. В состоянии покоя мышечного волокна Са2+-АТФаза саркоплазматиче- ского ретикулума активно закачивает ионы Са2+ из саркоплазмы в его цис¬терны, где большинство ионов находится в обратимо связанном состоянии с белком кальсеквестрином. Для того чтобы вызвать сокращение, потенциал действия мышцы, вы- зваемый двигательным нервом, распространяется по сарколемме мышеч¬ной клетки и внутрь поперечных трубочек (t-система), открывая кальцие¬вые ионные каналы мембраны. Далее происходит выход ионов Са2+ в саркоплазму. Когда концентрация ионов Са2+ в межфибриллярном пространстве, т. е. в саркоплазме между нитями актина и миозина, увеличивается до 10 мкМ/л (1(Г5 М) и более, ионы Са2+ начинают связываться с тропонином С. Активация тропонина С при связывании с ионами Са2+ вызывает смещение тропонин-тропомиози- нового комплекса примерно на 25° вокруг поверхности актинового фила¬мента и погружение молекул тропомиозина вглубь между белковыми цепя¬ми актина. В результате на актиновых филаментах открываются места свя¬зывания для головок поперечных мостиков миозина и начинается цикл их сокращения. При этом начинается сокращение всей скелетной мышцы, которое будет продолжаться до тех пор, пока концентрация ионов Са2+ в саркоплазме не станет пороговой или сверхпороговой для поддержания со¬кратительного процесса. 16. Виды и режимы сокращения мышц. Тетанус, виды тетануса, условия их возникновения. 1. Быстрое сокращение и расслабление скелетной мышцы в лабораторных условиях в ответ на стимуляцию называется одиночным мышечным сокращением (ОМС) – 1ый вид. Суммация ОМС при увеличении частоты раздражения приводит к тетаническому сокращению – 2ой вид. 2. Сокращение отдельного мышечного волокна подчиняется закону «все или ничего». 3. Амплитуда сокращения целой мышцы не подчиняется закону «все или ничего» и может увеличиваться при увеличении частоты и силы раздражения. 4. Амплитуда сокращения также увеличивается при увеличении количества мышечных волокон, участвующих в сокращении. При сокращении всех мышечных волокон амплитуда сокращения максимальная (оптимум). При дальнейшем увеличении частоты и силы раздражения амплитуда сокращения снижается – пессимум. 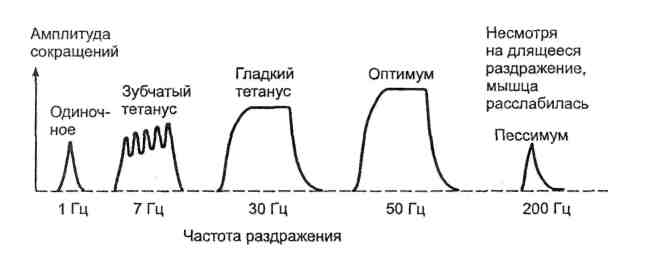 5. При невысокой частоте стимуляции каждое последующее раздражение попадает в фазу расслабления ОМС, и поэтому суммация ОМС будет неполной - зубчатый тетанус. При более высокой частоте стимуляции каждое последующее раздражение попадает в фазу сокращения ОМС, мышца не успевает расслабиться - гладкий тетанус. 5. При невысокой частоте стимуляции каждое последующее раздражение попадает в фазу расслабления ОМС, и поэтому суммация ОМС будет неполной - зубчатый тетанус. При более высокой частоте стимуляции каждое последующее раздражение попадает в фазу сокращения ОМС, мышца не успевает расслабиться - гладкий тетанус. 6. Режимы сокращения мышцы. Сокращение называется изометрическим (сопротивление давлению), если напряжение мышцы растет, но укорочения не происходит. Если мышца укорачивается при неизменном напряжении, то такое сокращение называется изотоническим (поднятие груза). Ауксотоническое – сокращение, при котором меняется и напряжение, и длина мышцы. |