Гиста 2022. Эпителиальная, ткани внутренней среды, мышечная и нервная ткани
 Скачать 5.1 Mb. Скачать 5.1 Mb.
|

|
Быков: Регенерация (обновление) эпидермиса обеспечивает его барьерную функцию благодаря постоянной замене и удалению наружных слоев, повреждающихся и содержащих микроорганизмы на своей поверхности. Период обновления равен 20-90 сут. (в зависимости от области тела и возраста); он резко сокращается при воздействии на кожу раздражающих факторов и при некоторых заболеваниях (например, псориазе). Эпидермальная пролиферативная единица (ЭПЕ) - самообновляющаяся единила эпидермиса, имеющая вид шестиугольной клеточной колонки, равной по ширине роговой чешуйке, а по высоте - толщине эпидермиса и включающая все его слои. Источником самообновления и поддержания структуры ЭПЕ служит деление базальных клеток, лежащих в ее основании вокруг 1-2 центрально расположенных стволовых клеток эпидермиса (по некоторым данным, в центре ЭПЕ лежит клетка Лангерганса - см. ниже). Базальные клетки далее мигрируют в колонку, в которой они вертикально перемешаются и дифференцируются, превращаясь в конечном итоге в роговые чешуйки. ЭПЕ описаны лишь в участках кожи с низкой скоростью обновления. Клетки Лангерганса (внутриэпидермальные макрофаги) - захватывают антигены, проникающие в эпидермис, осуществляют их процессинг и транспорт в лимфатические узлы, представляя лимфоцитам и вызывая развитие иммунной реакции. Имеют костномозговое происхождение, лежат в базальном или шиповатом слоях, содержат развитые органеллы и особые мембранные гранулы (Бирбека) в форме теннисной ракетки (функция неясна). Кузнецов:156. Кожа. Потовые железы: классификация, локализация, строение. Гормональная регуляция, иннервация.По Кузнецову 1. В коже имеются два главных компонента: – эпидермис — многослойный плоский ороговевающий эпителий – и лежащая под ним дерма — соединительнотканная основа. Под дермой обычно располагается подкожная жировая клетчатка, или гиподерма. Кроме того, в коже содержатся железы — потовые и сальные. Производными кожи являются также молочные железы — видоизмененные потовые железы. Существуют еще придатки кожи — волосы и ногти. 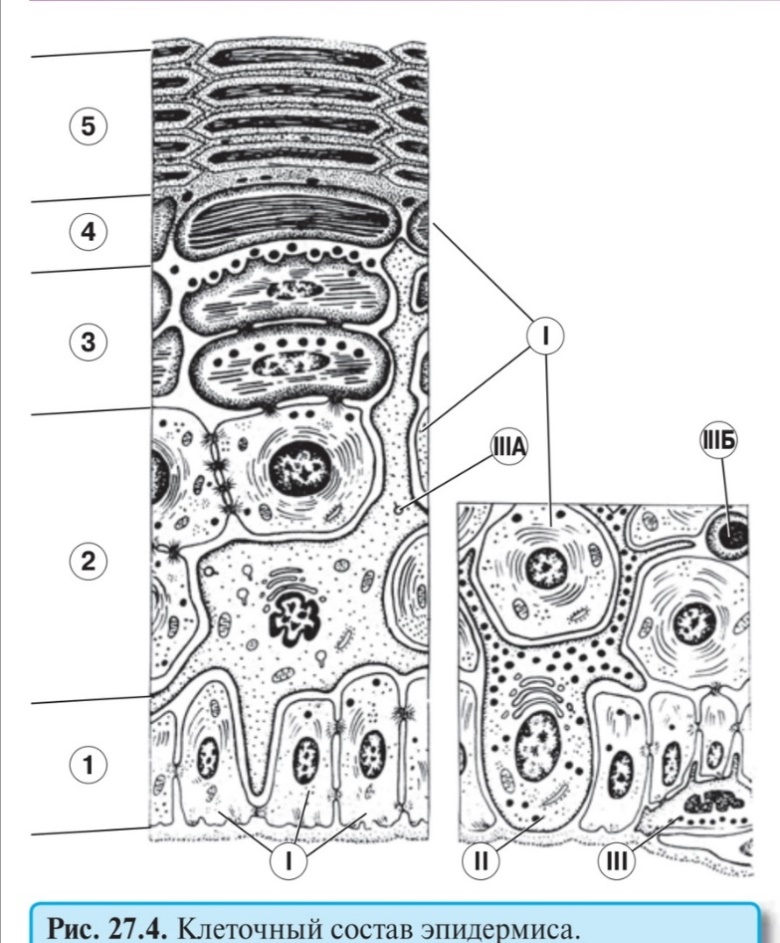 Эпидермис как «тонкой», так и «толстой» кожи содержит клетки следующих типов: кератиноциты (I), меланоциты (II), внутриэпителиальные макрофаги (клетки Лангерганса) (IIIА) и контактирующие с их отростками Т-лимфоциты (IIIБ), осязательные клетки Меркеля (IV), а также недавно обнаруженные клетки Гринстейна (на рисунке не показаны). Из них только кератиноциты располагаются во всех слоях эпидермиса — базальном (1), шиповатом (2), зернистом (3), блестящем (4) и роговом (5) Кератиноциты— основной вид клеток эпидермиса: они составляют не менее 85 % его клеток. Их общее название обусловлено высоким (и все возрастающим от слоя к слою) содержанием кератиновых тонофиламентов. Вместе с тем для кератиноцитов каждого слоя используется специальное обозначение: соответственно 1) базальные, 2) шиповатые и 3) зернистые кератиноциты, 4) кератиноциты блестящего слоя и, наконец, 5) роговые чешуйки, или корнеоциты. Меланоциты (II на рис. 27.4) имеют невральное происхождение: выселяются в эмбриогенезе из закладки нервной системы. Они располагаются в базальном слое эпидермиса, составляя 10–25 % клеток этого слоя. Десмосомных контактов с соседними клетками не образуют. По форме меланоциты — многоотростчатые. Главная их особенность — это наличие и в теле, и в отростках специфических мембранных органелл — меланосом, содержащих пигмент меланин. Меланосомы а) В меланосомах меланин синтезируется из аминокислоты тирозина (под действием ферментов тирозиназы и ДОФА-оксидазы) и накапливается в виде плотных гранул. б) У людей разной расы (разного цвета кожи) различно не количество меланоцитов в эпидермисе, а лишь количество и размер меланосом в клетках. Так, у темнокожих людей в меланоцитах много крупных меланосом с высоким содержанием меланина. Соответственно, у светлокожих людей количество меланоцитов такое же, но в них очень мало меланосом и меланина. Поэтому идентифицировать меланоциты на препаратах кожи в этом случае трудно. в) Из отростков меланоцитов эпидермиса меланосомы могут выходить в межклеточное пространство и оттуда фагоцитироваться другими клетками (не способными к синтезу меланина): кератиноцитами и макрофагами эпидермиса, а также меланоцитами дермы. г) Меланин, поглощая УФ-лучи, защищает подлежащие ткани. При высокой интенсивности солнечного облучения в меланоцитах эпидермиса компенсаторно увеличивается синтез меланина, что внешне воспринимается как загар. Клетки Лангерганса (внутриэпителиальные макрофаги) и клетки Гринстейна Клетки Лангерганса (IIIA на рис. 27.4), как и прочие макрофаги, происходят из моноцитов. В коже они располагаются в базальном и шиповатом слоях эпидермиса. И, подобно меланоцитам, не образуют десмосомных контактов с соседними клетками. Форма клеток — многоотростчатая (в эпидермисе отростки доходят до зернистого слоя), а ядро — многолопастное. Клетки Гринстейна Они тоже являются многоотростчатыми. Наиболее распространена версия, что – клетки Гринстейна представляют антигены Т-супрессорам, тогда как клетки Лангерганса представляют антигены Т-хелперам. И таким образом соблюдается баланс иммунологического ответа.Клетки Гринстейна имеют непосредственное отношение к иммунным реакциям в коже. Осязательные клетки Меркеля – один из видов механорецепторов кожи. Они отвечают (вместе с тельцами Мейснера) за тактильную чувствительность, или за осязание.Клетки Меркеля (1) находятся в базальном слое эпидермиса (особенно много их в кончиках пальцев) и в волосяных луковицах.Вероятно, они, как и меланоциты, имеют невральное происхождение. Клетки Меркеля могут располагаться поодиночке, но часто объединены в группы (до 20 клеток) — т.н. диски Меркеля. С соседними кератиноцитами клетки Меркеля образуют десмосомные контакты (чем отличаются от меланоцитов и клеток Лангерганса). К самим клеткам Меркеля подходят окончания дендритов (2) чувствительных нейронов. Тактильное раздражение клеток Меркеля генерирует нервный сигнал в этих окончаниях. Кроме того, клетки Меркеля содержат гранулы с гормоноподобными факторами: энкефалином, бомбезином, вазоинтестинальным пептидом (ВИП) и др. Эти вещества выделяются после тактильного раздражения клеток Меркеля, распространяются паракринным способом (т. е. путем диффузии в окружающем межклеточном пространстве) и влияют на регенерацию эпителия и тонус близлежащих кровеносных сосудов. Следовательно, с помощью тактильных воздействий можно стимулировать процессы заживления. Дерма «тонкой» кожи.  В связи с этой таблицей сделаем три замечания. а) Меланоциты дермы не синтезируют меланин, а, видимо, получают меланосомы от меланоцитов эпидермиса. Встречаются они лишь в определенных местах — в области анального отверстия и в околососковых кружках. б) Гладкие миоциты образуют в дерме не только мышцы, поднимающие волос, но и пучки, не связанные с волосами. Сокращение тех и других пучков на холоде приводит к появлению «гусиной кожи». Это сопровождается сжатием просвета близлежащих мелких сосудов, в результате чего уменьшаются и теплопотери. Потовые железы имеются и в «толстой», и в «тонкой» коже. За сутки они образуют 600–700 мл пота.В последнем 98 % массы составляет вода, а 2 % — растворенные компоненты: соли (NaCl и др.), продукты метаболизма (лактат, мочевина, мочевая кислота, аммиак и др.), некоторые белки. Функциональное значение потоотделения состоит в том, что это один из важнейших способов теплоотдачи и дополнительный способ выделения продуктов обмена. Большинство потовых желез стимулируется симпатической нервной системой, поэтому при волнении потоотделение усиливается. Однако эффекторным медиатором в симпатических окончаниях является в данном случае ацетилхолин (поскольку именно он обычно стимулирует секрецию желез в организме). Потовые железы — простые неразветвленные трубчатые. Их концевые отделы располагаются в глубине сетчатого слоя дермы — на границе с подкожной жировой клетчаткой — и представляют собой длинные трубочки, свернутые в клубочек. В этих отделах содержатся секреторные клетки— высокие, с округлыми ядрами, лежащие в один слой. Они делятся на два типа: – светлые – продуцирующие воду и ионы; – темные – секретирующие органические продукты метаболизма. На периферии концевых отделов находятся миоэпителиальные клетки, имеющие уплощенную форму. Своими сокращениями они способствуют выделению секрета. Скорее всего, именно на них действуют симпатические нервные волокна, стимулирующие потоотделение. Выводные протоки образованы двухслойным кубическим эпителием и представляют собой длинные трубочки, имеющие спиралеобразный ход. В своих начальных участках выводные протоки отличаются от концевых отделов меньшим диаметром и большей базофилией. Миновав дерму, выводной проток в большинстве случаев переходит в эпидермальную свою часть и открывается на поверхности потовой порой. При повреждениях кожи клетки выводных протоков потовых желез могут участвовать в регенерации эпидермиса. И в «тонкой» коже потовые железы обычно имеют такую же морфологию. По способу секреции потовые железы делятся на мерокриновые (эккриновые) и апокриновые. 1. Мерокриновые железы находятся в большей части кожи.При выделении секрета клетки сохраняют свою целостность.Отчасти поэтому содержание органических веществ в поте — низкое (около 2 %).Концевые отделы — небольшого размера (30–35 мкм).Выводные протоки открываются на поверхности кожи потовыми порами.Развиваются железы из эпидермальных почек, врастающих в мезенхиму (дерму). 2. Апокриновые железы содержатся в коже лба, подмышечных впадин и аногенитальной области.Выделение секрета сопровождается разрушением апикальных отделов секреторных клеток.Поэтому в секрете — относительно много (до 5 %) органических веществ (в т. ч. белков секреторных клеток). При разложении этих веществ на поверхности кожи возникает характерный резкий запах. Эфферентные окончания вегетативной нервной системы иннервируют в коже а) гладкие миоциты сосудов, б) внесосудистые гладкие миоциты и в) потовые железы. Кроме того, в коже много чувствительных нервных окончаний. 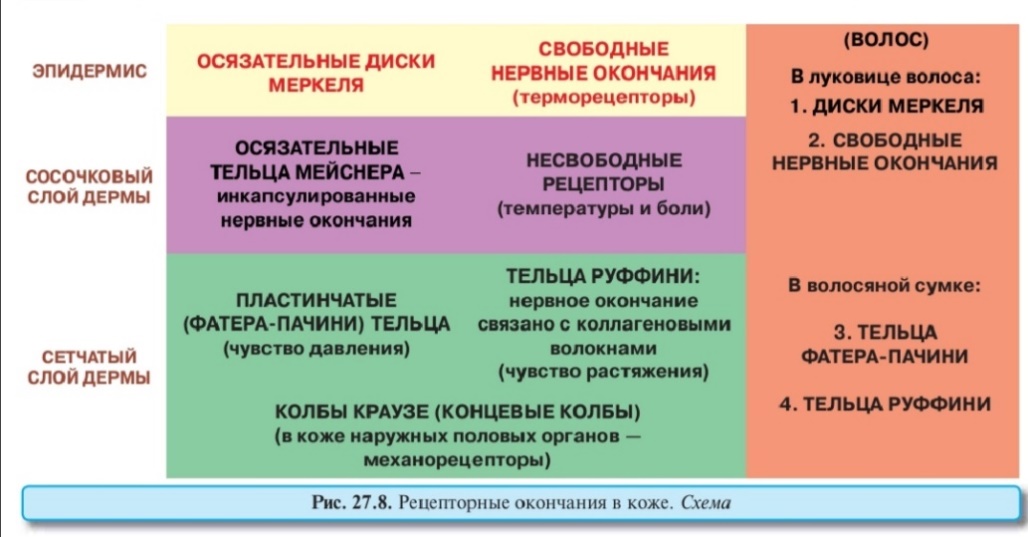 1. В эпидермисе имеются свободные нервные окончания: а) одни просто проникают между клетками эпителия и выступают в основном в качестве температурных и болевых рецепторов, б) а другие контактируют с осязательными дисками Меркеля. 2. В сосочковом слое дермы располагаются еще два вида рецепторов. а) Неинкапсулированные (и несвободные) нервные окончания, тоже воспринимающие температуру и боль. Термин «несвободные» означает, что конечные ветвления дендритов чувствительных нейронов сопровождаются леммоцитами. б) Второй тип рецепторов — осязательные тельца Мейснера, представляющие собой инкапсулированные нервные окончания. Каждое такое тельце включает уже три компонента: А) конечные ветвления дендрита, Б) окружающие их видоизмененные леммоциты и В) тонкую соединительнотканную капсулу. 3. Сетчатый слой дермы. Наиболее распространены пластинчатые тельца, или тельца Фатера—Пачини. Их главная особенность — очень толстая капсула, состоящая из фиброзных пластинок, разделенных тонкими слоями жидкости. Считается, что благодаря такой структуре рецептор воспринимает не только статическое давление, но и его быстрое изменение — вибрацию. Еще один вид рецепторов — тельца Руффини; их особенно много в коже подошвы. В этих тельцах окончания дендритов связаны с коллагеновыми волокнами и реагируют на их растяжение. В коже наружных половых органов содержатся также колбы Краузе. Здесь окончания дендритов очень сильно ветвятся, отчего окружающая их капсула имеет вид колбы. Все они являются инкапсулированными. Большинство вышеперечисленных видов рецепторов находится и во внутрикожных частях волос: а) в эпителиальных компонентах – осязательные диски Меркеля и свободные нервные окончания; б) а в соединительнотканной волосяной сумке – маленькие тельца Фатера-Пачини и тельца Руффини. 157. Волос. Строение волосяного фолликула, волосяной луковицы. Рост волоса. Пигментацияволоса. Иннервация волосяного фолликула 1. Волосы, как и связанные с ними сальные железы, распространены почти по всей поверхности «тонкой» кожи (за исключением ряда мест). Всего волос на голове и теле — несколько сотен тысяч. Длина волос — от нескольких миллиметров до 1,5–2,0 м. Скорость роста волоса — 0,3–0,4 мм/сут. I. При этом андрогены ускоряют рост волос на лице (усы, борода). II. Цитокины, влияя на пролиферацию эпителия, тоже стимулируют рост волос: так, последние быстро растут по краям заживающей раны. Различают три вида волос: а) длинные (волосы волосистой части головы, бороды, усов, подмышечных впадин и лобка), б) щетинистые (волосы бровей и ресниц), в) пушковые (остальные волосы). Волос любого типа образует комплекс с рядом структур дермы. Он включает: собственно волос, волосяной фолликул – эпителиальный мешок вокруг внутрикожной части волоса; дермальное влагалище – соединительнотканный мешок вокруг волосяного фолликула; дополнительные структуры – сальные железы и мышцу, поднимающую волос. Волосяной фолликул – это футляр (или мешок) эпителиальной природы, в который заключены луковица и корень волоса. Фолликул включает: – внутреннее эпителиальное влагалище, заканчивающееся на уровне дна волосяной воронки; – наружное эпителиальное влагалище, которое на уровне дна волосяной воронки переходит в эпидермис. Следует отметить, что под волосяным фолликулом нередко понимают не только пару эпителиальных влагалищ, но и содержащиеся внутри них луковицу и корень волоса. Волосяная луковица — самая нижняя часть волоса. Она контактирует с соединительнотканным волосяным сосочком и получает от него питание. Состоит в основном из ранних кератиноцитов: базальных (прилегают к волосяному сосочку) и шиповатых. Последние лежат многослойным пластом и, как обычно, имеют полигональную форму и крупное овальное ядро. Кроме того, в луковице содержатся все другие клетки, которые встречаются в нижних слоях эпидермиса: меланоциты, внутриэпителиальные макрофаги (клетки Лангерганса), осязательные клетки Меркеля Базальные и (отчасти) шиповатые кератиноциты волосяной луковицы активно делятся (каждые 1–3 суток), т. е. играют ту же роль, что и базальные клетки эпидермиса. Поэтому они называются матрицей волоса. Размножаясь, клетки матрицы дают начало всем трем слоям волоса (кутикуле, корковому и мозговому веществу) и внутреннему эпителиальному влагалищу. Волос в своем жизненном цикле проходит три фазы: фазу роста (фазу анагена),фазу инволюции сосочка и луковицы (фазу катагена),фазу покоя (фазу телогена) — до выпадения волоса. 1. Фаза катагена (инволюции) продолжается 2–4 недели. Она начинается с атрофии волосяного сосочка и, следовательно, с ухудшения питания волоса. Одними из первых на это реагируют меланоциты: они перестают образовывать пигмент; поэтому луковица волоса оказывается непигментированной. Затем прекращаются деления в матрице волоса. Уже существующие клетки волосяной луковицы завершают ороговение и (поскольку не вытесняются новыми клетками) остаются в луковице; так что последняя вместе с корнем (тоже полностью ороговевшим) превращается в роговую волосяную колбу. Внутреннее эпителиальное влагалище разрушается и исчезает (путем слущивания клеток). Наружное же эпителиальное влагалище сохраняется, и волосяная колба (вместе с волосом) перемещается по нему вверх, отчего нижняя часть влагалища запустевает. Важное обстоятельство: в основании наружного влагалища в норме должны оставаться стволовые клетки для волосяной луковицы нового волоса. 2–3. Фазы телогена старого волоса и анагена нового волоса. Непигментированная волосяная колба старого волоса пребывает в фолликуле (состоящем теперь только из наружного эпителиального влагалища) 2–4 месяца. Это — фаза покоя (фаза телогена) данного волоса. В конце данного периода на дне фолликула активизируются стволовые клетки: они формируют конусовидную луковицу нового волоса. Это стимулирует появление и нового сосочка. В результате начинаются рост нового волоса (фаза анагена) и образование внутреннего эпителиального влагалища. Через некоторое время растущий волос вытесняет из фолликула старый волос и появляется на поверхности кожи. (Заметим, что старый волос может выпасть и до этого.) Но это — только начало фазы анагена нового волоса. Полная ее продолжительность — до 2–6 лет. Все это время матрица волоса образует новые генерации клеток самого волоса и внутреннего эпителиального влагалища, что обеспечивает постоянный рост волоса. *В Кузнецове нет, поэтому из готового дока Пигментация волоса зависит от присутствия меланина, вырабатываемого меланоцитами в волосяной луковице. Отношения меланоцитов и кератиноцитов такие же как в эпидермисе. По мере дифференцировки кератиноциты захватывают меланин, который определяет пигментацию мозгового и коркового вещества. Волос человека иннервируется 5-9 нервными волокнами, которые идут ниже устья сальной железы, далее меняют направление и циркулярно обхватывают волос. От циркулярно идующих ответвляются волокна, проходящие параллельно стержню волоса в составе волосяной сумки выше расширения волосяной луковицы, образуя терминальный комлекс-палисадный (частокольный) аппарат. 1. Палисадный аппарат. Терминали контактируют со стекловидной мембраной и имеют форму уплощенного цилиндра, расположенного между шванновскими клетками. 2. Комплекс пило-Руффини. Представлен большим количеством афферентных волокон, ветвящихся по кольцу вокруг волосяного фолликула ниже устья сальной железы. Среди их терминалей имеются пучки коллагеновых волокон и соединительнотканные клетки-аналоги клеток периневрия, не имеющие собственной базальной мембраны и образующие подобие капсулы комплекса. 3. Клетки Меркеля. Компактные лентовидные скопления клеток проходят по окружности в эпителии волосяного фолликула вблизи волосяной воронки. 4. Мышца, поднимающая волос. Иннервируется симпатическими волокнами. 158-159. Дифферон: понятие Нейросекреторные клетки гипоталамуса. Локализация, гормоны. Пути транспорта гормонов. Ядра и нейросекреторные клетки гипоталамуса. Гормоны гипоталамуса. Дифферон — это совокупность клеточных форм (от стволовой клетки до высокодифференцированных), составляющих определенную линию дифференцировки. В ткани могут присутствовать клетки нескольких разных дифферонов. Причем, у взрослого человека одни диффероны представлены всеми своими клетками, а другие — только специализированными клетками (без предшествующих клеточных форм). Нейросекреторные клетки гипоталамуса 1) В группу крупноклеточных ядер гипоталамуса входят: – супраоптические ядра, лежащие над зрительным перекрестом, – паравентрикулярные ядра, расположенные в боковых стенках III желудочка мозга. В этих ядрах преобладают крупные нейросекреторные клетки. Они реагируют на холинергические раздражения со стороны вышележащих центров — лимбической системы, гиппокампа и др. В ответ на указанные раздражения (и, возможно, под влиянием и других сигналов) в клетках образуются два пептидных гормона: АДГ (вазопрессин) и окситоцин. Аксоны нейросекреторных клеток спускаются в заднюю долю гипофиза и образуют здесь на капиллярах аксовазальные синапсы. АДГ и окситоцин достигают по аксонам синапсов и попадают в кровь большого круга кровообращения. Место выхода нейрогормонов в кровь называют нейрогемальным органом. Следовательно, в данном случае таким органом является задняя доля гипофиза. В целом же крупноклеточные ядра гипоталамуса и задняя доля гипофиза составляют вместе т. н. гипоталамо-нейрогипофизарную систему. 2) B группу мелкоклеточных ядер гипоталамуса входят аркуатные, вентромедиальные и дорсомедиальные ядра. Они располагаются в средней части серого бугра и образуют аркуатновентромедиальный комплекс. В ядрах комплекса содержатся мелкие нейросекреторные клетки. Они реагируют, в частности, на адренергические и дофаминергические сигналы со стороны вышележащих центров. В клетках комплекса синтезируются гормоны, влияющие на клетки аденогипофиза, — либерины и статины. В данном случае аксоны нейросекреторных клеток идут в медиальное возвышение на нижней поверхности гипоталамической области и образуют здесь аксовазальные синапсы с первичными капиллярами портальной системы гипофиза. Эти капилляры являются фенестрированными. А базальная мембрана их эндотелиоцитов расщеплена на два листка, между которыми имеется перикапиллярное пространство. Окончания аксонов прилегают к наружному листку мембраны. Спустившиеся по аксонам либерины и статины вначале попадают в перикапиллярное пространство и лишь затем — в просвет капилляров. Капилляры собираются в портальные вены. По этим венам гормоны достигают передней и средней долей гипофиза, где влияют на их функционирование. Итак, мелкоклеточные ядра гипоталамуса и аденогипофиз составляют вместе единую функциональную систему - гипоталамо-аденогипофизарную. Лекция Валлиулина: Гипоталамо-гипофизарная система: Секреторными нейронами гипоталамус синтезирует и по аксонам доставляет в область срединного возвышения гормоны гликопротеиды – либерины (рилизинг-гормоны), которые секретируются в первичную капиллярную сеть (чудесную) и током крови доставляются в аденогипофиз (передняя и промежуточная доли), где вступают в контакт с его клетками. В гипоталамо-гипофизарной системе реализуется принцип обратной связи, при котром конечная концентрация гормонов крови ркегулилирует выработку факторов, приводящую к появлению этого гормона. Бывает как «-« , так и «+» В ядрах гипоталамуса синтезируются 2 гормона- белка, которые по аксонам доставляются в заднюю долю гипофиза (нейрогипофиз), где через аксо-вазальные синапсы секрет в области накопительных телец Херринга попадает в кровь, - окситоцин, вазопрессин (АДГ). Окситоцин (гормон родов): 1) вызывает сокращение ГМК, вазоконстриктор 2) стимулирует работу корзинчатых клеток молочных желез, способствуя лактации. Вазопрессин: 1) вазоконстрикция 2) усиливает канальцевую реабсорбцию натрия и воды в собирательных трубочках, в нефроне, что приводит к повышению АД. 160. Окситоцин. Синтез, транспорт, секреция, мишени, эффекты. По Кузнецову Синтезируется крупными нейросекреторными клетками крупноклеточных ядер (супраоптическое и паравентрикулярное) гипоталамуса. Содержащие продукты трансляции мембранные пузырьки транспортируются по аксонам этих нейронов в составе гипоталамо-нейрогипофизарного пути в заднюю долю гипофиза, и через аксо-вазальные синапсы гормоны секретируются в кровь. |