Физиология нервов, синапсов, мышц и рецепторов
 Скачать 23.71 Mb. Скачать 23.71 Mb.
|

|
Виды и механизмы памяти; мгновенная, кратковременная и долговременная память, и опыты их доказывающие. Механизм образования условных рефлексов, схема, ее иллюстрирующая. “Замыкание” временной связи - (Э.А.Асратян). Память – способность организма приобретать, сохранять и формировать в сознании информацию и навыки. Различают врожденную – генетическую и приобретенную – иммунную и нейронную память. Классификация памяти. По формам восприятия инфы различают слуховую и моторную; по уровням освоения – воспроизводящую и облегчающую; по длительности хранения информации - кратковременную и долговременную. Выделяют также ассоциативную и неассоциативную, процедурную и декларативную. Процедурная связана с усвоением навыков. Декларативная – хранение любой инфы (кроме навыков): формул, символов, содержание прочитанной книги. Хранение инфы в долговременной памяти называется энграммой. Процесс перехода инфы из электрофизиологической памяти в структурную носит название консолидации (промежуточная память). В этот период происходит образование энграмм. Выделяют произвольную, непроизвольную, образную и логическую. Механизм формирования памяти: на первом этапе происходят электрофизиологические процессы, на втором биохимические реакции, а на третьем структурные изменения в стуктурах и синапсах ЦНС. Кратковременная память – элекрофизиологическая, промежуточная – нейрохимическая, долговременная – нейроструктурная. Следует выделить первичную и вторичную электрофизиологическую память. Первичная электрофизиологическая память. Эта память кратковременна. Объем составляет 7+- единицы. Длительность хранения инфы составляет от нескольких секунд до 5 минут. В основе механизма первичной электрофизиологической памяти лежит кратковременная циркуляция возбуждения благодаря кратковременной синаптической потенциации. Начальной стадией усвоения любой инфы является восприятие раздражителя. Инфа воспринимается рецепторами органов чувств и кодируется в виде рецепторного потенциала, который с помощью своего электрического поля обеспечивает возникновение нервных импульсов, предающихся в ЦНС. Вторичная электрофизиологическая память. Основой механизма этого вида памяти является долговременная циркуляция возбуждения благодаря долговременной синаптической потенциации. Одним из факторов, активирующим этот вид памяти являются эмоции. Длительность хранения информации определяется длительностью циркуляции возбуждения (10 – 15 часов). Нейрохимическая (промежуточная) память. В данный период происходит консолидация памяти – перевод хранения инфы с помощью биохимических процессов в долговременную память. Консолидация продолжается не более четырех часов. Нейроструктурная (долговременная) память. Ее основой является синтез белка и она не нарушается при экстремальных воздействиях на мозг. Формируется с помощью электрофизиологических и нейрохимических процессов. На данном этапе формирования памяти можно видеть структурные изменения в синапсах и отростках нейронов (ультраструктурная память), происходят незаметные молекулярные структурные изменения в нейронах (макромолекулярная память). Образование условных рефлексов. Условные рефлексы формируются не только на уровне неокортекса, но и на уровне мозжечка. Предполагают, что мозжечок контролирует точность выполнения движений во времени, т.к. только он обладает способностью в любой момент времени заблокировать любую двигательную реакцию или, наоборот, дать ей возможность реализоваться. Основные правила выработки условных рефлексов. Необходимые условия:
Способствующие факторы:
Основные стадии образования условного рефлекса.
Механизм образования условного рефлекса (временной связи).
В начале выработки условного рефлекса временная связь образуется только за счет механизмов электрофизиологической памяти – распространения возбуждения между двумя возбужденными корковыми центрами, возбуждаемыми в результате действия безусловного и условного раздражителей. Образованию функциональной связи между корковыми центрами ассоциируемых раздражителей способствует синхронизация биопотенциалов. По мере действия условного и безусловного раздражителей и повторяющегося возбуждения соответствующих центров кратковременная память переходит в долговременную, происходят структурные изменения в нейронах.
Образование условных рефлексов с прямыми и обратными связями является общей закономерностью ВНД. В процессе выработки условного рефлекса образуется двусторонняя связь между двумя центрами: корковым концом сенсорной системы, на рецепторы которого действует условный раздражитель, и центром безусловного рефлекса, на базе которого вырабатывается условный рефлекс.
В начале выработки условного рефлекса наблюдается дивергенция возбуждения в ЦНС, затем развивается синаптическая потенциация на пути между двумя центрами (проторение пути) и, наконец, формируются доминантные отношения между двумя центрами. 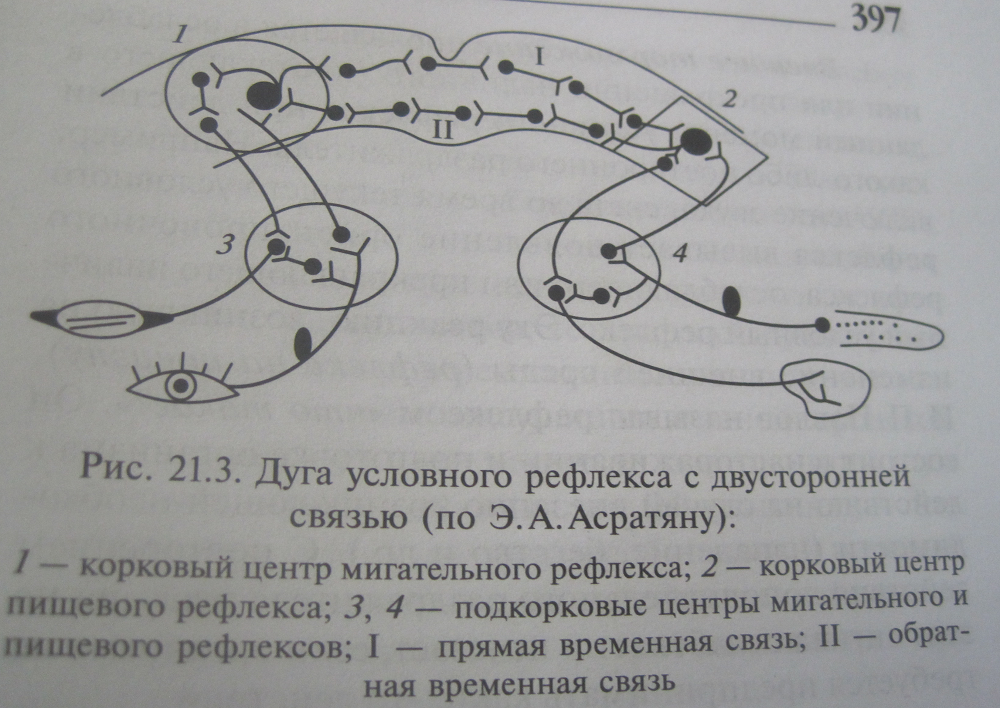
Торможение условных рефлексов. Различают врожденное и приобретенное торможение условных рефлексов. Безусловное торможение подразделяется на два вида внешнее и запредельное.
Механизм внешнего торможения: посторонний сигнал сопровождается появлением в коре большого мозга нового очага возбуждения, который при достаточной силе раздражителя оказывает угнетающее влияние на текущую условно – рефлекторную деятельность по механизму доминанты. Новый очаг возбуждения находится вне дуги наличного условного рефлекса. Значение внешнего раздражения состоит в том, что оно способствует экстренному приспособлению организма к меняющимся условиям внешней и внутренней среды и дает возможность при необходимости переключиться на другую деятельность в соответствии с ситуацией.
Механизм запредельного торможения: между силой условного торможения и величиной ответной реакции имеется определенное соответствие – закон силы: чем сильнее условный сигнал, тем сильнее условно – рефлекторная реакция. Выполняет охранительную функцию. Условное торможение. Локализовано в пределах нервного центра данного условного рефлекса. Различают четыре основных виды: угасательное, запаздывательное, дифференцировочное и условный тормоз.
Условное торможение можно ослабить и восстановить угасший ранее рефлекс, если во время проявления торможения подействовать посторонним сигналом – возникает ориентировочно – исследовательский рефлекс. Это явление называется растормаживанием. Механизм условного торможения условных рефлексов изучен недостаточно. На клеточном уровне зарегистрирована гиперполяризации корковых нейронов. В процесс условного торможения широко вовлекаются различные мозговые структуры, при угасательном торможении амплитуда ВПСП постепенно снижается без заметного перехода в гиперполяризацию клеточной мембраны. Торможение условных рефлексов можно объяснить тем, что при неподкреплении условного раздражителя доминантные взаимоотношения двух очагов возбуждения исключаются и остается только один очаг возбуждения, вызванный условным сигналом. Проторенный путь между доминантными очагами возбуждения постепенно устраняется, т.к. исчезает синаптическая потенциация, сформированная ранее. Значение всех видов условного торможения заключается в устранении ненужной в данное время деятельности.
Основные правила выработки условных рефлексов. Необходимые условия: 1. неоднократное повторение во времени действия индифферентного раздражителя с безусловным; 2.условный стимул должен предшествовать безусловному. Способствующие факторы: 1. Оптимальное состояние организма; 2. Активное функциональное состояние нервного центра безусловного рефлекса; 3. Отсутствие посторонних сигналов из внешней среды и от внутренних органов. 4. Оптимальное соотношение силы условного и безусловного раздражителей. В процессе образования условного рефлекса выделяют три стадии. Латентная стадия характеризуется выраженной концентрацией возбуждения в проекционных зонах коры условного и безусловного раздражителей и отсутствием условных поведенческих реакций. Стадия генерализации условного рефлекса состоит в том, что условные реакции возникают на сигнальный и другие раздражители, а также в интервалах между предъявлениями условного стимула. В этот период различные биоэлектрические сдвиги широко распространены по коре большого мозга и подкорковым структурам. Наблюдается синхронизация биоэлектрической активности во многих участках коры и подкорковых образованиях – дистантная синхронизация биопотенциалов, в основе которой лежит процесс дивергенции возбуждения. Стадия специализации обеспечивает дифференцировку стимулов, специализацию условно – рефлекторного навыка. Множество раздражителей внешнего мира и внутренней среды организма воспринимаются рецепторами и становятся источниками импульсов, которые поступают в кору больших полушарий. Здесь они анализируются, различаются и синтезируются, соединяются, обобщаются. Способность коры разделять, вычленять и различать отдельные раздражения, дифференцировать их и есть проявление аналитической деятельности коры головного мозга. Сначала раздражения анализируются в рецепторах, которые специализируются на световых, звуковых раздражителях и т. п. Высшие формы анализа осуществляются в коре больших полушарий. Аналитическая деятельность коры головного мозга неразрывно связана с ее синтетической деятельностью, выражающейся в объединении, обобщении возбуждения, которое возникает в различных ее участках под действием многочисленных раздражителей. В качестве примера синтетической деятельности коры больших полушарий можно привести образование временной связи, которое лежит в основе выработки условного рефлекса. Сложная синтетическая деятельность проявляется в образовании рефлексов второго, третьего и высших порядков. В основе обобщения лежит процесс иррадиации возбуждения. Анализ и синтез связаны между собой, и в коре происходит сложная аналитико-синтетическая деятельность. Динамический стереотип. Внешний мир действует на организм не единичными раздражителями, а обычно системой одновременных и последовательных раздражителей. Если система последовательных раздражителей часто повторяется, это ведет к образованию системности, или динамического стереотипа в деятельности коры головного мозга. Таким образом, динамический стереотип представляет собой последовательную цепь условно-рефлекторных актов, осуществляющихся в строго определенном, закрепленном во времени порядке и являющихся следствием сложной системной реакции организма на сложную систему положительных (подкрепляемых) и отрицательных (неподкрепляемых, или тормозных) условных раздражителей. Условно-рефлекторное переключение. Понятие "условно-рефлекторное переключение" ввел Э.А.Асратян. Оно состоит в том, что один и тот же раздражитель может стать условным сигналом различных условных реакций. Физиологическим механизмом условно-рефлекторного переключения является образование комплексов разного сигнального значения из одних и тех же основных сигналов с добавлением разных обстановочных раздражителей. Например, гудок в одной экспериментальной камере может быть сигналом пищевой реакции, а в другой камере - сигналом оборонительного рефлекса, или, один и тот же сигнал в первую половину дня может служить оборонительным условным раздражителем, а во вторую половину - пищевым сигналом. Из этого примера видно, что здесь имеет место образование разных условных рефлексов на комплексы раздражителей, общим компонентом которых служит сигнал, применяемый экспериментатором в качестве условного раздражителя (гудок). Мозг животного во время опыта анализирует составные части комплекса раздражителей, выделяет "основные" агенты и "переключатели". Он синтезирует, объединяет их и связывает с определенной деятельностью.
Критерии оценки свойств НС. Сила процесса возбуждения отражает работоспособность нервной клетки. Она проявляется в функциональной выносливости, т.е. способности выдерживать длительное или кратковременное, но сильное возбуждение, не переходя в состояние охранительного торможения. Если слабое раздражение вызывает сильно возбуждение, ведущее к развитию охранительного торможения, то сила процесса возбуждения слабая, а тип НС высоковозбудимый. Сила процесса торможения понимается как функциональная работоспособность НС при реализации условного торможения. Сила процесса торможения проявляется в способности к образованию различных тормозных условных реакций – угасания, дифференцировки или запаздывания. Уравновешенность означает равенство силы процессов возбуждения и торможения, вовлеченных в выработку условных рефлексов или условного торможения. Подвижность нервных процессов указывает на способность НС быстро реагировать на изменения среды и переходить от одних условных рефлексов к другим в зависимости от требований среды. Павлов выделил 4 основных типа ВНД: 1. Сильный, уравновешенный, подвижный; 2. Сильный, неуравновешенный; 3. Сильный, уравновешенный, инертный; 4. Слабый. Сильный, уравновешенный, подвижный = сангвиник. Такой человек решительный, энергичный, с высокой возбудимостью, подвижен. Сильный, уравновешенный, инертный = флегматик. Спокойный, медлительный, со слабы проявлением чувств. Сильный, неуравновешенный = холерик. Вспыльчивый, с высоким уровнем активности, энергичный. Слабый = меланхолик. Унылый, тоскливый, склонный к мрачным мыслям. У человека тип ВНД определяют, во – первых, посредством выработки различных условных рефлексов и условного торможения; во – вторых с помощью специальных методик. Существует два методических подхода определения силы нервных процессов: исследования изменения реакций при разной интенсивности раздражителя и при длительном раздражении. В последнем случае сила определяется кА устойчивость величины латентного периода сенсомоторной реакции на многократное повторение раздражителей средней величины. Один из вариантов заключается в следующем: испытуемому дается 100 световых раздражителей с интервалом 1 – 3 с, каждый раз при загорании лампочки испытуемый должен как можно сильнее нажать на рычаг. Показателем, характеризующим силы нервных процессов, будет среднее значение самой двигательной реакции на последние 20 раздражений. Определение уравновешенности нервных процессов чаще всего проводится по количеству положительных двигательных реакций на тормозный раздражитель при выработке дифференцировки. Для этого испытуемому дается инструкция, не смотря на циферблат, останавливать стрелку на цифре 10. Высчитывают число точных, опережающих и опаздывающих реакций, вычисляют среднюю величину отклонений времени, вариационный размах и алгебраическую сумму отклонений. Для исследования подвижности нервных процессов используют показатели длительности последействия условного тормоза на двигательные реакции, соотношение латентных периодов условных двигательных реакций до и после переделки стереотипа. Испытуемому дается задача при загорании зеленой лампочки нажимать рычажок вправо, при загорании красной влево. Подают 50 раздражителей. Затем дают обратную задачу. Также подают 50 раздражителей. Количественными характеристиками подвижности являются среднее время реакции и точность. Роль генотипа в формировании ВНД. Доказательства влияния наследственного фактора получены в различных опытах. Так, в опыте Рундквиста от популяции активных крыс через 12 поколений получились активные крысы, а от пассивных пассивные. Влияние среды показано в опытах на щенках Майорова и Выржиковского. Одна группа животных содержалась в закрытых клетках, а вторая группа в свободных вольерах. У собак первой группы выработалось сильное пассивно – оборонительное поведение. |