Глик Молекулярная биотехнология. Глик Б., Пастернак Дж. Молекулярная биотехнология. Принципы и применение. Пер с англ. М. Мир, 2002. 589 с
 Скачать 9.74 Mb. Скачать 9.74 Mb.
|

Аттенуированные вакциныВ некоторых случаях в качестве живых вакцин можно использовать генетически модифицированные (рекомбинантные) микроорганизмы (бактерии или вирусы). Такие вакцины содержат либо непатогенные микроорганизмы, синтезирующие антигенные детерминанты определенного патогенного агента, либо штаммы патогенных микроорганизмов, у которых модифицированы или делетированы гены вирулентности, В этих случаях основные антигенные детерминанты являются составными компонентами бактериальных или вирусных частиц и имеют такую же конформацию, какую они принимают в болезнетворном микроорганизме. Изолированный же антиген часто утрачивает исходную конформацию и вызывает лишь слабый иммунный ответ. Вакцины 235
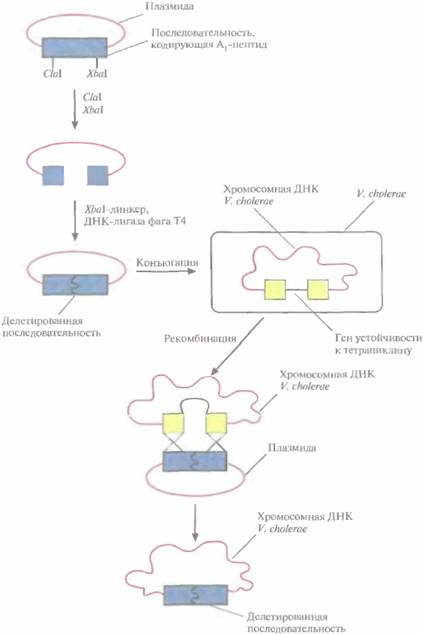
Противохолерные вакциныЖивые вакцины, как правило, гораздо более эффективны, чем неживые или субъединичные. Основное требование, предъявляемое к ним, -отсутствие в инокуляционном материале вирулентных микроорганизмов. Это требование учитывалось и при создании живой противохолерной вакцины. Холера — быстро развивающаяся кишечная инфекция, характеризующаяся лихорадкой, диареей, болью в животе, дегидратацией; передается через питьевую воду, загрязненную фекалиями. В развивающихся странах, где системы очистки воды и удаления сточных вод недостаточно развиты, угроза холеры вполне реальна. Возбудителем холеры является Vibriochoierae. Бактерия размножается в тонком кишечнике и выделяет в большом количестве энтеротоксин, который и ответствен за патогенный эффект. Энтеротоксин — это гексамерный белок: он состоит из одной субъединицы А, которая обладает ADP-рибозилирующей активностью и стимулирует аденилатциклазу, и пяти субъединиц В, которые специфически связываются с клеточным рецептором слизистой кишечника. Субъединица А имеет два функциональных домена: А1, обладающий токсической активностью, и А2, отвечающий за ее связывание с субъединицами В. В настоящее время используется противохолерная вакцина, содержащая убитые фенолом холерные вибрионы; она обеспечивает только частичную защиту от инфекции и лишь в течение 3—6 мес. Поэтому были предприняты попытки создать другие типы противохолерной вакцины. Как показали проведенные ранее исследования, субъединичная вакцина, содержащая инак-тивированный холерный энтеротоксин, не приводит к выработке полноценного иммунитета. 236 ГЛАВА 11 Поскольку V. cholerae колонизирует слизистую кишечника, разумно было предположить, что наиболее эффективной будет пероральная противохолерная вакцина. Имея это в виду, создали штамм V. cholerae, из генома которого была делетирована часть кодирующей А1-пептид нуклеотидной последовательности. Этот штамм не синтезирует энтеротоксин, а потому не является патогенным и подходит для создания живой вакцины. Эксперимент состоял в следующем. В ген А1-пептида V. choleraeбыл встроен ген устойчивости к тетрациклину. При этом прерывалась рамка считывания для Α1-пептида, но штамм становился устойчивым к тетрациклину. Его нельзя было использовать в качестве вакцины и потому, что со временем происходила спонтанная утрата тетрациклинового гена, и синтез энтеротоксина восстанавливался. Чтобы обойти эту проблему, создали штамм с дефектной нуклеотидной последовательностью, кодирующей А1-пептид, которая не могла восстанавливаться (рис, 11.6). Для этого использовали следующий подход, 1. Плазмиду, которая содержала сегмент ДНК, кодирующий А,-пептид, обработали рестри-цирующими эндонуклеазами С1аIи XbaI, каждая из которых расщепляла только кодирующую a1-пептид последовательность вставки. 2. Чтобы замкнуть плазмиду в кольцо, к ClaI-сайту пришили ХbаI-линкер и обработали плазмиду рестриктазой XbaI. 3. С помощью ДНК-лигазы фага Т4 соединили XbaI-сайты плазмиды. В результате из середины кодирующей А1-пептид последовательности оказался удаленным сегмент длиной 550 п. н., соответствующий аминокислотным остаткам 183-194. 4. С помощью конъюгации перенесли эту плазмиду в штамм V. cholerae, несущий ген устойчивости к тетрациклину в локусе, кодирующем Α1-пептид. 5. В результате рекомбинации между оставшейся в составе плазмиды частью последовательности, кодирующей А1-пептид, и геном А,-пептида, прерванным Теtг-геном, кодирующая А1-пептид последовательность в хромосоме была замешена гомологичным плазмидным сегментом с делецией. 6. Внехромосомная плазмида не могла долго существовать в холерном вибрионе и через несколько поколений была утрачена. 7. Отобрали клетки с интегрированной дефектной А1 -кодирующей последовательностью, используя их чувствительность к тетрациклину. Полученный таким способом стабильный штамм с делегированной кодирующей А1-пептид последовательностью не синтезировал активный энтеротоксин и при этом сохранял все остальные биохимические свойства патогенной формы V. cholerae. Проводимые в настоящее время клинические испытания эффективности этой формы как противохолерной вакцины пока не дали однозначного результата. Вакцина обеспечивает почти 90%-ную защиту от холеры, но у некоторых испытуемых наблюдаются побочные эффекты. Возможно, понадобится изменить другой хромосомный локус этого штамма, чтобы можно было использовать его как вакцину. Противосальмонеллезные вакциныДругой способ получения непатогенных штаммов, пригодных для создания на их основе живых вакцин, состоит в удалении из генома патогенных бактерий хромосомных областей, отвечающих за независимые жизненноважные функции. При этом лучше делетировагь по крайней мере две такие области, поскольку вероятность их одновременного восстановления очень мала. Предполагается, что штамм с двойной делецией будет обладать ограниченной пролиферативной способностью и сниженной патогенностью, но обеспечит выработку иммунного ответа. Разные штаммы Salmonellaвызывают острые кишечные инфекции, постнагальную инфекцию, брюшной тиф, пищевую токсикоинфек-цию. Для профилактики всех этих заболеваний совершенно необходимо иметь эффективную вакцину. Чтобы получить аттенуированные штаммы Salmonella, вносили делеции в гены ara, кодирующие ферменты биосинтеза ароматических соединений, и в гены pur, кодирующие ферменты метаболизма пуринов. Такие штаммы с двойной делецией вызывают легкую форму инфекции и обладают в 106 раз меньшей вирулентностью. На их основе уже созданы эффективные Вакцины 237
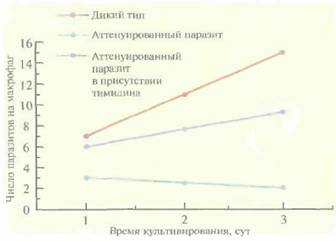
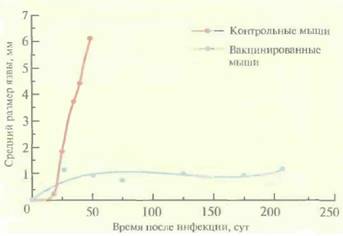
пероральные вакцины для мышей, овец, крупного рогатого скота, цыплят, а совсем недавно — и для человека. Противолейшманиозные вакциныПростейшие паразиты Leishmaniaтоже могут вызывать у человека развитие иммунного ответа, однако создание эффективных вакцин против них представляет собой нелегкую задачу. С этой целью можно использовать аттенуированные линии Leishmania, но они часто ревертируют и становятся вирулентными, а кроме того, могут длительное время бессимптомно персистировать в организме человека — резервуаре инфекции - и передаваться другим людям. Чтобы решить эти проблемы, попытались создать атте- 238 ГЛАВА 11 нуированную неревертирующую линию Leishmaniaс помощью делеции одного из важных для метаболизма генов (например, гена дигидрофолатредуктазы—тимидилатсинтазы). У одного из таких паразитов, LeishmaniamajorЕ10-5АЗ, два гена дигидрофолатредуктазы—тимидилатсинтазы были заменены генами устойчивости к антибиотикам G-418 и гигромицину. В отличие от паразитов дикого типа, при выращивании L. majorE10-5A3 в обычной культуре или в культуре макрофагов в среду необходимо добавлять тимидин (рис. 11.7). В организме мышей BALB/c паразиты оставались жизнеспособными в течение нескольких дней, что достаточно для создания стойкого иммунитета у животных (рис. 11.8), но недостаточно для развития заболевания. Ни персистирующая инфек-
ция, ни заболевание не возникали даже у наиболее чувствительных видов мышей, так что данная линия вполне подходит для создания вакцины. Проведя дополнительные эксперименты на животных, можно будет проверить ее эффективность при иммунизации человека. «Векторные» вакциныПротивовирусные вакциныВ качестве эффективной живой противооспенной вакцины широко используют вирус коровьей оспы (ВКО), относящийся к роду поксвирусов. Геном этого вируса полностью секвенирован; он представляет собой двухцепочечную ДНК длиной 187 т.п.н., кодирующую примерно 200 разных белков. ДНК ВКО реплициру-
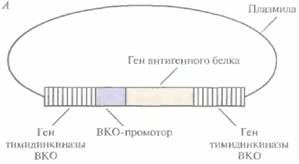
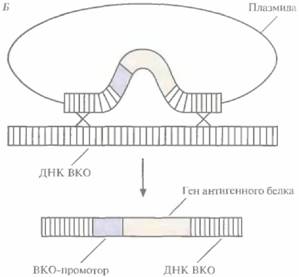
Вакцины 239 ется в цитоплазме инфицированных клеток, а не в ядре, благодаря наличию у вируса генов ДНК-полимеразы, РНК-полимеразы и ферментов, осуществляющих кэпирование, метилирование и полиаденилирование мРНК. Поэтому, если в геном ВКО встроить чужеродный ген, так чтобы он находился под контролем ВКО-промотора, то он будет экспрессироваться независимо от регуляторных и ферментных систем хозяина. ВКО имеет широкий спектр хозяев (позвоночных и беспозвоночных), остается жизнеспособным в течение многих лет после лиофилизации (испарения воды с помощью замораживания) и не обладает онкогенными свойствами, а потому может использоваться для создания так называемых векторных вакцин. С их помощью осуществляется доставка и экспрессия в организме-хозяине клонированных генов, кодирующих антигенные белки, которые индуцируют выработку протективных антител. Геном ВКО имеет большие размеры и не содержит уникальных сайтов рестрикции, что не позволяет встраивать в него дополнительные нуклеотидные последовательности. Однако нужные гены можно вводить в геном ВКО с помощью гомологичной рекомбинации in vivo следующим образом. 1. Сегмент ДНК, кодирующий специфичный антиген (например, HBcAg), встраивают в плaзмидный вектор непосредственно после клонированного ВКО-промотора, включенного в какой-либо несущественный ген ВКО, например ген тимидинкиназы (рис. 11.9, А). 2. Этой плазмидой трансформируют культуру дефектных по тимидинкиназе животных клеток, обычно фибробластов куриного эмбриона, предварительно инфицированных ВКО дикого типа, который синтезирует функциональную тимидинкиназу, 3. В результате рекомбинации между нуклеотидными последовательностями, фланкирующими промотор и ген протективного антигена, и гомологичными последовательностями вирусного генома происходит встраивание клонированного гена в вирусную ДНК (рис. 11.9, Б). Частота таких рекомбинаций невысока, однако популяцию клеток, содержащих реком-
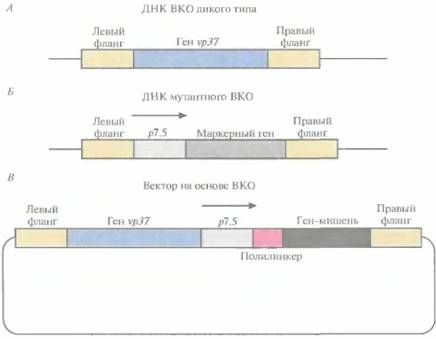
бинантный ВКО, можно обогатить, используя селективную среду с бромдезоксиуридином. Этот токсичный аналог тимидина в отсутствие тимидинкиназы не включается в синтезируемую ДНК и не оказывает токсического действия. Дефектные по тимидинкиназе клетки-хозяева, которые содержат обычный ВКО, в присутствии бромдезоксиуридина погибают, а клетки, несущие рекомбинантный ВКО с разрывом в гене ти- 240 ГЛАВА 11 мидинкиназы, становятся устойчивыми к его токсическому действию. 4. Проводят окончательный отбор с помощью ДНК-зонда, гибридизующегося с геном антигенного белка. Поскольку дефектные по тимидинкиназе ВКО спонтанно возникают с относительно высокой частотой (примерно 1 на 103—104 вирусных частиц), нередко проводят котрансфекцию клеток каким-либо селективным маркером и нужным геном. Это облегчает разграничение спонтанных мутантов и мутантов, полученных с помощью гомологичной рекомбинации. В качестве селективного маркера обычно используют ген пео, кодирующий фермент неомицин-фосфотрансферазу [I и обеспечивающий устойчивость к аналогу канамицина G-418. Этот ген, в отличие от других селективных маркеров, остается стабильным при встраивании в геном ВКО. Разработана специальная система, позволяющая избежать прерывания рамки считывания генов ВКО при встраивании чужеродного гена. При этом отпадает необходимость в использовании селективных маркеров, поскольку каждый образующий бляшку рекомбинантный вирус будет содержать и экспрессировать ген-мишень. ДНК ВКО дикого типа несет ген νρ37, отвечающий за образование бляшек при росте вируса в монослойной культуре животных клеток (рис. 11.10, А). Если заменить этот ген маркерным геном Е. соli, то образуется мутантный ВКО, который не формирует бляшки при выращивании его в течение 2—3 сут в культуре животных клеток (рис. 11.10, Б). Ген-мишень вводят в этот мутантный вирус с помощью гомологичной рекомбинации его ДНК с вектором, несущим ген νρ37 и ген-мишень (рис. 11.10, В). Мутантный ВКО, получивший ген νρ37, приобретает способность к образованию бляшек, при этом в его геном встраивается ген-мишень, а маркерный ген утрачивается. Мутантный вирус с делегированным геном vp37 не может ревертировать к дикому типу, поэтому каждая вирусная частица, образующая бляшку, содержит желаемую конструкцию. Этот метод прост, применим для переноса и экспрессии любого гена-мишени, не требует каких-либо дополнительных маркерных генов и не прерывает рамку считывания генов ВКО. В геном ВКО уже удалось встроить и экспрессировать в культуре животных клеток несколько генов антигенных белков: G-белка ви-
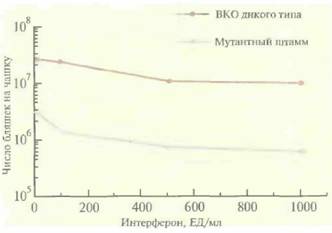
Вакцины 241 руса бешенства, поверхностного антигена гепатита В, поверхностных белков вируса Синдбис, NP- и НА-белков вируса гриппа, N- и G-белков вируса везикулярного стоматита, гликопротеинов вируса простого герпеса. Некоторые из полученных на основе ВКО рекомбинантных векторов можно использовать для создания эффективных вакцин. Так, рекомбинантный ВКО, экспрессирующий ген гликопротеина D вируса простого герпеса типа 1, предотвращает герпесные инфекции у мышей, а рекомбинантный ВКО, экспрессирующий ген поверхностного антигена вируса бешенства, индуцирует выработку протективных антител у лис, основных переносчиков вируса бешенства в Европе. Векторные ВКО-вакцины позволяют провести иммунизацию сразу от нескольких заболеваний. Для этого можно использовать рекомбинантный ВКО, который несет несколько генов, кодирующих разные антигены. В зависимости от используемого ВКО-промотора чужеродный белок может синтезироваться в ранней или поздней фазе инфекционного цикла, при этом его количество определяется силой промотора. Обычно для достижения высокого уровня экспрессии используют «поздние» ВКО-промоторы: p11 (промотор гена, отвечающего за синтез белка мол. массой 11 кДа) или рСАЕ (промотор гена интегрального белка вируса коровьей оспы типа А). При встраивании в одну ДНК ВКО нескольких чужеродных генов каждый из них помещают под контроль отдельного ВКО-промотора, чтобы предотвратить гомологичную рекомбинацию между различными участками вирусной ДНК, которая может привести к утрате встроенных генов. Живая рекомбинантная вирусная вакцина имеет ряд преимуществ перед неживыми вирусными и субъединичными вакцинами: 1) презентация аутентичного антигена практически не отличается от таковой при обычной инфекции; 2) вирус может реплицироваться в клетке-хозяине и увеличивать количество антигена, который активирует продукцию антител В-клетками (гуморальный иммунитет) и стимулирует выработку Т-клеток (клеточный иммунитет); 3) встраивание генов антигенных белков в один и большее число сайтов генома ВКО еще больше уменьшает его вирулентность. Недостаток живой рекомбинантной вирусной вакцины состоит в том, что при вакцинации лиц со сниженным иммунным статусом (например, больных СПИДом) у них может развиться тяжелая вирусная инфекция. Чтобы решить эту проблему, можно встроить в вирусный вектор ген, кодирующий человеческий интерлейкин-2, который стимулирует Т-клеточный ответ и ограничивает пролиферацию вируса. Нежелательные побочные эффекты пролиферации ВКО можно предупредить инактивацией вируса после вакцинации. Для этого был создан чувствительный к интерферону вирус (ВКО дикого типа относительно устойчив к его действию), пролиферацию которого можно регулировать в случае возникших при вакцинации осложнений. Механизм устойчивости ВКО к интерферону оставался неустановленным, пока не была обнаружена открытая рамка считывания K3L, кодирующая белок мол. массой 10,5 кДа. Этот белок содержит аминокислотную последовательность, гомологичную N-концевой части эукариотического фактора инициации elF-2α мол. массой 36,1 кДа. N-концевые области обоих белков содержат 87 практически идентичных аминокислотных остатков, причем в положении 51 в обоих случаях находится серии, который в elF-2α фосфорилируется активируемой интерфероном Р1-киназой, что приводит к ингибированию синтеза белка в обработанных интерфероном клетках. К3L-белок действует как конкурентный ингибитор фосфорилирования elF-2α, обеспечивая устойчивость ВКО к интерферону, и если из генома ВКО удалить ген K3L или его часть, то вирус станет чувствительным к интерферону, С помощью ΠЦР-мутагене за гена K3L, находящегося в составе плазмиды, и последующей гомологичной рекомбинации между ДНК ВКО и плазмидой с целью замены К3L-последовательности дикого типа модифицированным вариантом был сконструирован мутантный ВКО K3L . Этот штамм оказался в 10—15 раз более чувствительным к интерферону, чем штамм дикого типа (рис. 11.11). Эта работа является важным этапом на пути создания более безопасных ВКО-векторов. Последовательности, сходные с K3L, могут содержать и другие устойчивые к интерферону вирусы, что позволит с помощью де- 242 ГЛАВА 11
леций создавать их штаммы, чувствительные к интерферону. Большинство работ по созданию живых вирусных вакцин проводились на ВКО, однако в качестве кандидатов на роль векторов для вакцинации рассматриваются и другие вирусы: аденовирус, полиовирус и вирус ветряной оспы. Вектор на основе живого аттенуированного полиовируса (его исследования только начинаются) привлекателен тем, что позволяет проводить пероральную вакцинацию. Такие «слизистые» вакцины (вакцины, компоненты которых связываются с рецепторами, расположенными в легких или желудочно-кишечном тракте) пригодны для профилактики самых разных заболеваний: холеры, брюшного тифа, гриппа, пневмонии, мононуклеоза, бешенства, СПИДа, болезни Лайма. Но до любых клинических испытаний любого на первый взгляд безобидного вируса как системы доставки и экспрессии соответствующего гена необходимо убедиться в том, что он действительно безопасен. Например, повсеместно используемый ВКО вызывает у людей осложнения с частотой примерно 3,0-10-6. Поэтому из генома рекомбинантного вируса, который предполагается использовать для вакцинации человека, желательно удалить последовательности, ответственные за вирулентность. Противобактериальные вакциныДля лечения заболеваний, вызываемых бактериями, широко используются антибиотики, и лишь совсем недавно начались работы по созданию противобактериальных вакцин. Стимулом к этому стали весьма веские причины. • Не все бактериальные инфекции поддаются лечению антибиотиками. • Широкое использование антибиотиков в последние 40 лет привело к появлению большого числа устойчивых к ним бактериальных штаммов. • Во многих тропических странах отсутствуют условия для хранения антибиотиков. • Часто больные прекращают лечение раньше, чем это предписано врачом, или принимают антибиотики в недостаточной дозе. Имея в виду, что создаваемая противобактериальная вакцина должна быть достаточно эффективной, важно выбрать правильную стратегию. Если патогенная бактерия плохо растет в культуре и сложно получить ее аттенуированный штамм, необходимо использовать альтернативные способы. Например, Rickettsiarickettsii, грамотрицательная облигатная внутриклеточная бактерия, вызывающая пятнистую лихорадку Скалистых гор, не растет в культуре. Чтобы обойти эту трудность, создали субъединичную вакцину, содержащую основной поверхностный антиген R. rickettsii, белок мол. массой 155 кДа. Она эффективно защищала мышей от этой патогенной бактерии. Бактерии кок системы доставки антигеновАнтигены, находящиеся на наружной поверхности бактериальной клетки, обладают более высокой иммуногенностью, чем локализованные в цитоплазме. Поэтому один из подходов, исполь- Вакцины 243 зуемых при создании вакцин, состоит в размещении протективного антигена патогенной бактерии на поверхности живой непатогенной бактерии. Многие бактерии имеют жгутики, состоящие из белка флагеллина; под микроскопом они выглядят как нити, отходящие от бактериальной клетки. Если сделать так, что жгутики непатогенного микроорганизма будут нести специфический эпитоп патогенного микроорганизма, то можно будет индуцировать выработку протективных антител. Именно такой подход использовали при создании противохолерной вакцины. Синтетический олигонуклеотид, кодирующий эпитоп субъединицы В холерного токсина, встроили в гипервариабельный участок гена флагеллина Salmonellaи полученную конструкцию ввели в дефектный по флагеллину штамм Salmonella. При этом было известно, что эпитоп, включающий 50—64 аминокислотные остатки субъединицы В холерного токсина, индуцирует выработку антител к интактному холерному токсину. Химерный флагеллин нормально функционировал, а эпитоп холерного токсина размещался на поверхности жгутиков. Иммунизация мышей с помощью интраперитонеальной инъекции примерно 5-106 живых или убитых формалином бактерий с модифицированным флагеллином индуцировала выработку большого количества антител как к пептиду (50—64 аминокислотным остаткам), так и к молекуле интактного холерного токсина. Аналогичным образом можно встраивать два и даже три разных эпитопа в один флагеллиновый ген Salmonellaи создать поливалентную противобактериальную вакцину. С помощью перорального введения аттенуированных штаммов Salmonellaможно осуществлять доставку в организм хозяина многих бактериальных, вирусных и паразитарных антигенов. Большую роль при этом играет выбор промотора, контролирующего транскрипцию чужеродного гена. Если используется слишком сильный промотор, может возникнуть метаболическая «перегрузка», сдерживающая пролиферацию бактерий. В отличие от ферментера, организм животного-хозяина не является замкнутой системой, и экспрессию чужеродного гена нельзя регулировать изменением температуры или добавлением специфических метаболитов. Регуляторную роль может играть только промотор, реагирующий на те или иные сигналы. Например, работу промотора nirB E. coli можно регулировать, изменяя содержание нитритов и кислорода в среде, а наиболее активен он в анаэробных условиях. В одном из экспериментов промотор nirB использовали для контроля экспрессии гена нетоксичного иммуногенного С-фрагмента столбнячного токсина в аттенуированном штамме Salmonella. В развивающихся странах инфекция Clostridium tetani уносит более 1 млн. жизней в год. Если генетически модифицированный штамм Salmonellaвыращивать в аэробных условиях, то С-фрагмент столбнячного токсина синтезироваться не будет. При пероральном же введении этой бактерии тестируемым мышам С-фрагмент синтезируется, и у животных вырабатываются антитела к нему. Таким образом, штамм Salmonella, в который встроен С-фрагмент столбнячного токсина, находящийся под контролем промотора nirB, можно использовать как живую пероральную противостолбнячную вакцину. Чтобы выяснить, насколько эффективен этот подход при вакцинации человека, необходимо провести дополнительные исследования. ЗАКЛЮЧЕНИЕТрадиционные вакцины содержат инактивированные или аттенуированные патогенные микроорганизмы (бактерии или вирусы). Эти вакцины имеют ряд недостатков: не все патогенные микроорганизмы можно вырастить в необходимых для производства вакцины количествах; работа с большим количеством патогенных микроорганизмов требует соблюдения строжайших мер предосторожности; аттенуированные штаммы нередко ревертируют и становятся вирулентными; инактивация часто бывает неполной; срок годности вакцины зависит от условий ее хранения. Технология рекомбинантных ДНК позволяет создавать надежные вакцины, используя при этом разные подходы. Делетируя гены, ответственные за вирулентность, получают живые вакцины, содержащие непатогенные, иммунолога-чески активные штаммы, которые не могут 244 ГЛАВА 11 ревертировать и становиться патогенными. Клонированные гены, кодирующие основные антигенные детерминанты патогенного организма, встраивают в геном непатогенного носителя (обычно вируса) и получают безопасную, не содержащую болезнетворных микроорганизмов вакцину. Наконец, гены или их сегменты, кодирующие основные антигенные детерминанты патогенных микроорганизмов, встраивают в экспрессирующие векторы, получают нужный продукт в большом количестве и используют его как вакцину. Последний подход позволяет производить субъединичные и пептидные вакцины (если используются полноразмерные гены в первом случае и фрагменты генов, кодирующих домены основных антигенных детерминант — во втором). Пептидные вакцины получают и с помощью химического синтеза пептидов. ЛИТЕРАТУРАAndio R., D. Silvera, S. D. Suggett, P. L. Achacoso, C. J. Miller, D. Baltimore, M. B. Feinberg. 1994. Engineering poliovirus as a vaccine vector for the expression of diverse antigens. Science 265: 1448-1451. Bittle J. L., R. A, Houghten, H. Alexander, T. M. Shinnick, J. G. Sutcliffc, R. A. Lernen, D. J. Rowlands, F, Brown. 1982. Protection against foot-and-mouth disease by immunization with a chemically synthesized peptide predicted from the viral nucleotide sequence. Nature 298: 30-33. Blancou J., M. P. Kieny, R. Lathe, J. P. Lecoq, P. P. Pastoret, J. P. Soulebot, P. Desmettre. 1986. Oral vaccination of the fox against rabies using a live recombinant vaccinia vaccine. Nature 322: 373-375. Blasco R., B. Moss. 1995, Selection of recombinant vaccinia viruses on the basis of plaque formation. GeneW: 157-162. Boothroyd J. C., P. E. Highfîeld, G. A, M. Cross, D. J. Rowlands, P. A. Lowe, F. Brown, T. J. R. Harris. 1981. Molecular cloning of foot and mouth disease virus genome and nucleotide sequences in the structural protein genes. Nature 290: 800-802. Brown Г. 1984. Synthetic viral vaccines. Annu. Rev. A/kro&O/. 38:221-235. Brown F. 1985. Peptides as the next generation of foot-and-mouth disease vaccines. Bio/Techno-logy 3:445-448. Burnette W. N. 1990. The advent of recombinant pertussis vaccines. Bio/Technology 8: 1003-1005. Charles L, G. Dougan. 1990. Gene expression and the development of live enteric vaccines. Trends Biotechnol. 8:117-121. Chatfield S. N., I. G. Charles, A. J. Makoff, M. D. Oxer, G. Dougan, D. Pickard, D. Slater, N. F. Fairweather. 1992. Use of the nirB promoter to direct the slnble expression of heterologous antigens in Salmonella oral vaccine strains: development of a single-dose oral tetanus vaccine. Bio/Technology 10: 888-892. Chow M., R. Yabrov, J. Bittle, J. Hogle, D. Baltimore. 1985. Synthetic peptides from four separate regions of the poliovirus type 1 capsid protein VP1 induce neutralizing antibodies. Proc. Natl. Acad. Sei. USA 82: 910-914. Clarke B. E., S. E. Newton, A. R, Carroll, M. J. Francis, G. Appleyard, A. D. Syred, P. E. Highfield, D. J. Rowlands, F. Brown. 1987. Improved immunogenicity of a peptide epitope after fusion to hepatitis В core protein. Nature 330: 381-384. Cohen J. 1993. Naked DNA points way to vaccines. Science 259: 1691-1692. Cremer К. J., M. Macketl, С. Wohlenberg, A. L. Notkins, B. Moss. 1985. Vaccinia virus recombinant expressing herpes simplex virus type 1 glycoprotein D prevents latent herpes in mice. Science. 228: 737-740. DiMarchi R., G. Brooke, C. Gale, V. Cracknel), T. Doel, N. Mowat. 1986. Protection of cattle against foot-and-mouth disease by a synthetic peptide. Science 232: 639-641. Ferguson M. 1991. Progress towards rabies control. Trends Biotechnol. 9: 7—11. Finkelstein A., R. F. Silva. 1989. Live recombinant vaccines for poultry. Trends Biotechnol. 7: 273-277. Flexner C., A. Hugin, B. Moss. 1987. Prevention of vaccinia virus infection in immunodeficient mice by vector-directed IL-2 expression. Nature 330: 759 262 Вакцины 245 Graham F. L. 1990. Adenoviruses as expression vector and recombinant vaccines. Trends Biotechnoi. 8: 85-87. Horwitz M. A., B.-W. E. Lee, B. J. Dillon, G. Harth, 1995. Protective immunity against tuberculosis induced by vaccination with major extracellular proteins of Mycobacterium tuberculosis. Proc. Natl. Acad. Sei. USA 92: 1530-1534. Jones T. R., S. L. Hoffman. 1994. Malaria) vaccine development. Clin. Microbioi. Rev. 7: 303—310. Kaper J. B., H. Lockman, M. M. Baldini, M. M. Levine. 1984. A recombinant live oral choiera vaccine. Bio/Technology 2: 345-349. Kaper J. В., H. Lockman, M. M. Baldini, M. M. Levine. 1984. Recombinant nontoxinogenic Vibrio choier-ae strains as attenuated cholera vaccine candidates. Nature 308: 655-658. Kaper J. В., J. G. Morris, Jr., M. M. Levine. 1995. Choiera. Clin. Microbioi. Rev. 8: 48-86. Kaslow D. C., S. N. Isaacs L A. Quakyi, R. W. Gwadz, B, Moss, D. B. Keister. 1991. Induction of Plasmodium falciparum transmission-blocking antibodies by recombinant vaccinia virus. Science 252: 1310-1313. Кupper H., W. Keller, С. Kurz, S. Forss, H. Schauer, R. Franze, K. Strohmaier, O. Marquardt, V. G. Zaslavsky, P. H. Hofschneidtr. 1981. Cloning of cDNA of major antigen of fool and mouth disease virus and expression in E. colt. Nature 289: 555-559. Lasky L. A., D. Dowbenko, C. C. Simonsen, P. W. Berman. 1984. Protection of mice from lethal herpes simplex virus infection by vaccination with a secreted form of cloned glycoprotein D. Bio/Technology 2: 527-532. Lowe R. S., P. M. Keller, B..!. Ketch, A. J. Uavison, Y. Whang, A. J. Morgan, E. KicfT, R. W. Elfe. 1987. Varicella-zoster virus as a live vector for the expression of foreign genes, Proc. Natl. Acad. ScL USA 84: 3896-3900. Mekalanos J. J., J. C. Sadoff. 1994. Cholera vaccines: fighting an ancient scourge. Science 265: 1387-1389. Michel M.-L., H. L. Davis, M. Schleef, M. Mancini, P. Tiollais, R. G. Whalen. 1995. DNA-mediated immunization to the hepatitis В surface antigen in mice: aspects of the humoral response mimic hepatitis В viral infection in humans. Proc. Natl. Acad. Sei. USA 92: 5307-5311. Miner J. N., D. E. Hruby. 1990. Vaccinia virus: a versatile tool for molecular biologists. Trends Biotechnoi 8: 20-25. Moss B. 1991- Vaccinia virus: a tool for research and vaccine development, Science252: 1662—1667. Nabel G. J., P. L· Feigner. 1993. Direct gene transfer for immunotherapy and immunization. Tnends Biotechnoi. 11:211-215. Newton S. M. C., C. O. Jacob, B. A. U. Stocker. 1989. Immunoresponse to cholera toxin upitope inserted in Salmonella flagellin. Science 244: 70-72. Nussenzweig R. S., C. A. Long. 1994. Malaria vaccines: multiple targets. Science 265: 1381—1383. Oehen S., H. Hengartner, R. M. Zinkernagel. 1991. Vaccination for disease. Science 251: 195-197. Paoletti E., J. TarlagHa January 1995. Interferon sensitive recombinant poxvirus vaccine. U.S. patent 5, 378,457. Ratafia M. 1987. Worldwide opportunities in genetically engineered vaccines. Bio/Technology 5: N54-1158. Sizemore D, R., A. A. Branstrom, J. C. Sadoff. 1995, Attenuated Shigella as a DNA delivery vehicle for DNA-mediated immunization. Science 270: 299-302. Stover C. K., V. F. de la Cruz, T. R. Fuerst, J. E. Buriein, L· A. Benson, L.T Bennett, G. P. Bansal, J. F. Young, M. H. Lee, G. F. Hatfull, S. B. Snapper, R. G. Barletta, W. R. Jacobs, Jr., B. R. Bloom. 1991. New use of BCG for recombinant vaccines. Nature 351: 456—460. Tang D.-C-, M. DeVit, S. A. Johnston. 1992. Genetic immunization is a simple method for eliciting an immune response. Nature 356: 152-154. Tartaglia J., E. Paoletti. 1988. Recombinant vaccinia virus vaccines. Trends Biotechnoi. 6: 43-46. Titus R. GM J. G. Gueiros-Filtio, L. A. R. De Freitas, S. M. Beverly. 1995. Development of a safe live Leishmania vaccine line by gene replacement. Proc. Natl. Acad. Sei. USA92: 10267-10271. Uhner J. В., J. J. Donnelly, S. Parker, G. II. Rhodes, P. L. Feigner, V. J. Dwarki, S. H. Gromkowski, R. R. Deck, C. M, DeWïtt, A. Friedman, L. A. I lawe, K. R. Leander, D. Martine/, H. C. Perry, J. W. Shiver, D. I,. Montgomery, M. A. Liu. 1993. 246 ГЛАВА 11 Heterologous protection against influenza by injection of DN A encoding a viral protein. Science259: 1745-1749. КОНТРОЛЬНЫЕ ВОПРОСЫ1. Опишите вкратце способ создания вакцины против бактерий, продуцирующих токсин. 2. Что ограничивает применение традиционных вакцин? 3. Предположим, что вы принимаете участие в работе международной организации по охране здоровья животных и вам нужно создать вакцину против крайне вирулентного вируса крупного рогатого скота. Известно, что геном представляет собой полиаденилированную линейную одноцепочечную РНК длиной 10 т. п. н. и содержит восемь разных генов. Вирус не имеет оболочки, его основной антигенной детерминантой является белок капсида (VP 2). Какую стратегию вы используете? 4. Какие подходы применяются при создании пептидных противовирусных вакцин? 5. Что представляет собой вирус коровьей оспы и как с его помощью можно получать уникальные живые рекомбинантные вакцины? 6. Предположим, что вы выделили РНК-содержащий вирус, вызывающий бешенство у скунсов и енотов. Как на основе этого очищенного вируса создать рекомбинантную вакцину, защищающую животных от бешенства? 7. Как работник Всемирной организации здравоохранения вы должны найти оптимальный способ искоренения бешенства в популяциях диких животных. Предположим, что у вас есть пептидная вакцина и вакцина на основе вируса коровьей оспы; выберите одну из них и обоснуйте ваше решение. 8. Как можно использовать в качестве вакцины бактерии со жгутиками? 9. Перечислите преимущества живой рекомбинантной вирусной вакцины перед неживой и субъединичной вакцинами. 10. Опишите несколько подходов, использованных при создании холерной вакцины. |