Лекции биохимия 3. Лекция Обмен липидов
 Скачать 4.12 Mb. Скачать 4.12 Mb.
|

Нуклеотидный состав ДНК И РНКДНК РНК Сокращённые обозначения дезоксиадениловая адениловая кислота кислота (дАМФ) (АМФ) А дезоксигуаниловая гуаниловая кислота кислота (дГМФ) (ГМФ) Г дезоксицитидиловая цитидиловая кислота кислота (дЦМФ) (ЦМФ) Ц дезокситимидиловая - кислоты (дТМФ) Т - уридиловая кислота (УМФ) У ДНК содержится в вирусах, клетках низших организмов (прокариот) и в ядрах клеток высших организмов (эукариот), а также в митохондриях и хлоропластах высших организмов. Кроме того, в цитоплазме прокариот и эукариот найдены малые кольцевые ДНК. В составе растительной ДНК значительная часть цитозина в остатках нуклеотидов метилирована (до 25 %) и содержится в виде 5-метилцитозина, с участием которого образуется ещё одна разновидность нуклеотидов – 5-метилдезоксицитидиловая кислота. РНК в основном локализованы в цитоплазме клеток, где они являются главными факторами, инициирующими синтез ферментов и других белков. В составе многих растительных вирусов в качестве основного генетического материала содержится не ДНК, а РНК. Молекулы нуклеиновых кислот представляют собой длинные цепочки, состоящие из остатков нуклеотидов, которые соединены через остатки ортофосфорной кислоты фосфодиэфирными связями. В образовании фосфодиэфирных связей участвуют гидроксильные группы рибозы или 2-дезоксирибозы, связанные с третьим и пятым углеродными атомами. Однако на концах образуемой полинуклеотидной цепи остаются свободные гидроксильные группы у третьего и пятого углеродных атомов рибозы или дезоксирибозы, которые обозначают 3'-ОН и 5'-ОН. Соответствующие концы полинуклеотидной цепи, содержащие 3'-ОН и 5'-ОН, называют 3'- и 5'- концами. На прочность фосфоэфирных связей в составе полинуклеотидных цепей оказывают влияние углеводные компоненты образующих эти цепи нуклеотидов – рибоза или дезоксирибоза. Межнуклеотидные связи в молекулах РНК значительно лабильней по сравнению с ДНК, в связи с чем они легче подвергаются гидролизу. Последовательность соединения нуклеотидных остатков в молекулах нуклеиновых кислот фосфодиэфирными с 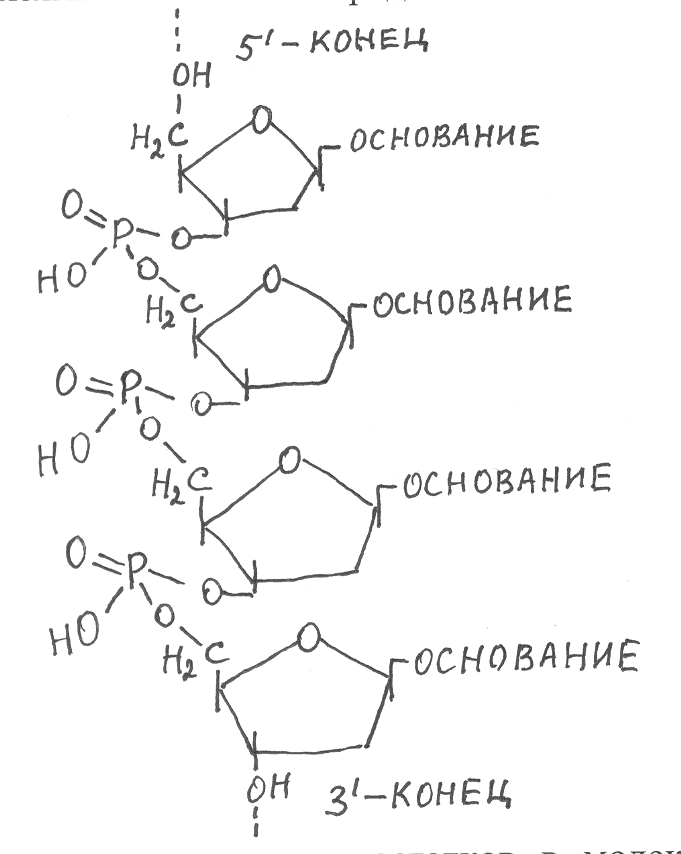 вязями может быть представлена в виде следующей схемы Число нуклеотидных остатков в молекулах нуклеиновых киcлот очень велико и оно варьирует в зависимости от вида организмов. Молекулы РНК могут включать от 100 до 100 тыс. нуклеотидных остатков (н.о). Ещё длиннее полинуклеотидные цепи ДНК: у вирусов – до 8-10 тыс. н.о., бактерий и других микроорганизмов – 106-107 н.о., у высших организмов – 108-1010 н.о. Чередование нуклеотидных остатков в полинуклеотидной цепи зависит от вида организмов. Учитывая, что она строится из четырёх типов нуклеотидов, может возникнуть огромное разнообразие макромолекул, общее число которых определяется по формуле N=4n, где n – общее количество нуклеотидных остатков в полинуклеотидной цепи. Например, если молекула нуклеиновой кислоты построена из 1000 нуклеотидов, то исходя из выше указанных условий может образоваться 41000 нуклеотидных последовательностей, каждая из которых представляет собой молекулу определённого генотипа. Генетическая роль и строение ДНКВпервые генетическая роль ДНК была установлена в опытах по изучению трансформации у бактерий. В 1928 г. Ф. Гриффит показал, что при введении в организм животных совместно с непатогенными штаммами пневмококков убитых нагреванием клеток патогенного штамма животные погибали от пневмонии. Однако если животным вводили отдельно непатогенный штамм или клетки убитого патогенного штамма, то заболевания пневмонией не наблюдалось. В связи с этим возникло предположение, что какое-то вещество, содержащееся в убитых клетках патогенного штамма, передаёт клеткам непатогенного штамма способность противостоять иммунной системе животных и поражать их пневмонией. Наблюдаемое явление назвали трансформацией бактерий. Для выявления трансформирующего фактора из убитых клеток были выделены в очищенном состоянии ДНК, белки и некоторые другие химические компоненты, которые стали вводить в организм животных совместно с клетками непатогенного штамма. Проводя такие опыты, О. Эйвери, К. Мак-Леод и М. Мак-Карти в 1944 г. установили, что трансформирующим фактором у пневмококков является ДНК. Таким образом, они впервые показали, что наследственная способность одной клетки осуществлять определённую биохимическую функцию может передаваться другой клетке путём введения в неё ДНК, выделенной из первой клетки. После этого в ряде других опытов с вирусами, бактериями и высшими организмами было подтверждено, что именно молекулы ДНК являются носителями генетической информации, определяющей наследственные свойства организмов. Открытие генетической роли ДНК послужило активным импульсом для дальнейшего изучения состава, строения и свойств нуклеиновых кислот. При изучении химического состава ДНК Э. Чаргаффом были выяснены закономерности нуклеотидного состава ДНК, которые получили название правила Чарграффа. Молекулы ДНК подвергали гидролизу, в ходе которого расщеплялись не только фосфодиэфирные связи, соединяющие остатки нуклеотидов, но и внутринуклеотидные связи между азотистым основанием и дезоксирибозой, а также между дезоксирибозой и остатком фосфорной кислоты. После гидролиза определяли содержание азотистых оснований. При этом было отмечено, что в составе ДНК любых организмов мольные соотношения пар оснований аденина и тимина, а также гуанина и цитозина всегда равны. Наблюдалось также равенство между суммой пуриновых и суммой пиримидиновых оснований, тогда как суммарное количество аденина и тимина заметно отличалось от суммарного содержания гуанина и цитозина. Используя сокращенные обозначения, отмеченные закономерности в содержании азотистых оснований в составе ДНК можно записать в виде следующих соотношений: А = Т; Г= Ц; А+Г = Т+Ц; А+Т≠ Г+Ц Изучение нуклеотидного состава ДНК различных организмов показало, что соотношение пар азотистых оснований А+Т/Г+Ц является важным показателем специфичности ДНК у разных организмов. Каждый вид организмов имеет свойственный ему нуклеотидный состав ДНК. В таблице 13 показано содержание азотистых оснований в ДНК животных, растений и микроорганизмов. 13. Содержание и соотношение азотистых оснований в ДНК различных организмов
Нуклеотидный состав ДНК у одного и того же генотипа остаётся постоянным в изменяющихся условиях окружающей среды, что является важнейшим фактором сохранения наследственных свойств организма. Кроме особенностей нуклеотидного состава, специфичность ДНК, свойственная конкретному генотипу, определяется также последовательностью соединения остатков нуклеотидов в полинуклеотидной цепи, которую называют первичной структурой ДНК. Каждый вид ДНК характеризуется совершенно определённой первичной структурой. Последовательность нуклеотидных остатков в полинуклеотидной цепи записывают в виде сокращённых обозначений нуклеотидов слева направо, начиная от 5'-конца. Буквенные обозначения нуклеотидов могут быть соединены чёрточками. Например, в нуклеотидной последовательности Т-Ц-Г-А-Ц-А слева направо последовательно соединены фосфодиэфирными связями остатки дТМФ, дЦМФ, дГМФ, дАМФ, дЦМФ и дАМФ, причём на 5'-конце данного олигонуклеотида находится остаток дТМФ, на 3'-конце – остаток дАМФ. Изучение вытянутых нитей ДНК методом рентгеноструктурного анализа выявило, что молекулы ДНК имеют пространственную структуру в виде спирали, включающей более одной полинуклеотидной цепи. А методом кислотно-щелочного титрования было показано, что нативная структура ДНК стабилизируется водородными связями. Учитывая эти данные, а также основываясь на правилах Чаргаффа, Д. Уотсон и Ф. Крик в 1953 г. разработали модель строения ДНК. Согласно модели Д. Уотсона и Ф. Крика молекула ДНК представляет собой двойную правозакрученную спираль, состоящую из двух полинуклеотидных цепей, которые закручены между собой и вокруг общей оси. Эти цепи антипараллельны и поэтому на каждом из концов двойной спирали находится 5'-конец одной и 3'-конец другой цепи. Сахарофосфатные группировки, входящие в состав нуклеотидов ДНК, выходят на поверхность спиралевидной структуры, тогда как азотистые основания находятся внутри и они соединяются водородными связями. При этом водородные связи возникают между парами оснований, которые структурно совместимы в пространстве. Такой принцип построения молекулы ДНК получил название комплементарного строения. В соответствии с принципом комплементарности одна полинуклеотидная цепь двойной спирали ДНК имеет последовательность соединения нуклеотидных остатков, которая закономерно отражает другую, антипараллельную полинуклеотидную цепь (рис. 41). Методом построения молекулярных моделей было выяснено, что пространственно совместимыми (комплементарными) являются пары азотистых оснований аденин – тимин и гуанин – цитозин, между которыми и возникают водородные связи, удерживающие вместе в двойной спирали две полинуклеотидные цепи. При этом между аденином и тимином возникают две водородные связи, а между гуанином и цитозином – три ( рис. 42). Один виток спирали включает по 10 нуклеотидов в каждой из двух цепей и имеет длину по оси спирали 3,4 нм, а межнуклеотидные расстояния составляют 0,34 нм. Диаметр двойной спирали ДНК – 1,8 нм. Плоскости оснований в двойной спирали ДНК перпендикулярны оси спирали, а соседние по направлению цепей пары оснований находятся под углом 36°. Как позднее было установлено, модель структуры ДНК Д. Уотсона и Ф. Крика отображает пространственное строение основной формы ДНК, в виде которой она существует в клетках живых организмов. Её назвали В-формой. В В-форме остаток дезоксирибозы имеет S-конформацию, а азотистые основания – анти-конформацию. Однако в дальнейшем было выяснено, что молекула ДНК может принимать и другую конформацию. Методом рентгеноструктурного анализа показано, что при понижении концентрации воды или добавлении спирта в ДНК изменяется пространственная структура дезоксирибозы, которая принимает N-конформацию. В результате перестройки структуры остатка дезоксирибозы уменьшаются расстояния между нуклеотидными остатками (до 0,25 нм), вследствие чего увеличивается диаметр двойной спирали, а число нуклеотидных пар в одном витке спирали возрастает до 11. Однако спираль остаётся правозакрученной и конформация азотистых оснований не изменяется. Двойную спираль, имеющую указанные параметры, называют А-формой ДНК. Между А-формой и В-формой ДНК существуют ещё и переходные варианты пространственной структуры двойной спирали. При рассмотрении строения ДНК следует отметить, что под влиянием внешней среды молекула ДНК может переходить из одной конформации в другую, но её нуклеотидный состав и последовательность соединения нуклеотидов остаются неизменными, что является важным условием для сохранения генетической информации, которая заложена, как мы увидим далее, именно в виде последовательности нуклеотидов. Для сохранения стабильности молекул ДНК важное значение имеет также принцип комплементарного связывания оснований в двойной спирали ДНК, в результате чего, зная последовательность соединения нуклеотидов в одной цепи, можно определить первичную структуру другой цепи, образующей с первой двойную спираль. Так, например, если в одной из цепей двойной спирали ДНК имеется последовательность нуклеотидных остатков -А-Г-Ц-Т-Ц-Г-, то совершенно очевидно, что в другой цепи согласно принципу комплементарности оснований образуется последовательность -Т-Ц-Г-А-Г-Ц-. Указанная закономерность в построении молекул ДНК предопределяет возможность синтеза новых молекул ДНК с сохранением заданной последовательности нуклеотидов, что осуществляется при размножении клеток организмов. У высших организмов молекулы ДНК локализованы главным образом в хромосомах клеточного ядра, по одной молекуле в каждой хромосоме. В связи с тем, что длина полинуклеотидной цепи ДНК во много раз (» 10 тыс. раз) превышает линейные размеры хромосом, она образует строго упорядоченные компактные структуры, которые принимают наименьшие линейные размеры в метафазных хромосомах. Основные структурные единицы при упаковке ДНК в хромосомах – нуклеосомы, которые формируются с участием гистоновых белков. Нуклеосома представляет собой октамер, содержащий по 2 молекулы каждого из четырёх гистонов – Н2А, Н2В, Н3 и Н4, на который закручена двойная спираль ДНК длиной 150-200 пар нуклеотидов. Диаметр нуклеосомы – 10 нм, толщина 5,7 нм. Образование нуклеосомы происходит в результате взаимодействия положительно заряженных аминокислотных остатков гистонов с отрицательно заряженными фосфатными группировками нуклеотидов ДНК, а также взаимодействия между гидрофобными группировками молекул гистонов, образующих гидрофобные глобулы. Между нуклеосомами находятся участки ДНК длиной 20-90 пар нуклеотидов в виде двойной спирали. Под воздействием физиологической среды нуклеосомы, соединённые свободными участками двойной спирали ДНК, формируют микрофибриллы диаметром 10 нм. Далее с участием гистонов Н1 и в присутствии катионов Са2+ микрофибриллы диаметром 10 нм закручиваются в левую суперспираль с диаметром 30 нм и превращаются в крупные хроматиновые фибриллы, которые, взаимодействуя со структурными белками хромосомы, образуют суперспирализованные петли, называемые доменами, закреплённые в структуре хромосомы в строго определённом месте. В среднем в составе одного домена обычно насчитывается 40-50 тыс. нуклеотидных пар, включающих группу функционально связанных генов. В опытах показано, что в упаковке хроматиновых фибрилл в составе метафазных хромосом принимают также участие гистоны Н1. В клетках прокариот (низших организмов) двойные спирали молекул ДНК образуют замкнутую кольцевую структуру, основой формирования которой служат белки негистоновой природы. Эта структура получила название нуклеоида. В составе нуклеоида белки негистоновой природы образуют комплексы с молекулой ДНК, переводя её в высокоупорядоченное состояние. У растений и других высших организмов не вся ДНК содержится в хромосомах ядра, часть её находится в виде кольцевых структур в составе митохондрий и пластид. Митохондриальная и пластидная ДНК содержит генетическую информацию, необходимую для осуществления этими органеллами клетки их биологических функций. Как и ДНК прокариот, кольцевые молекулы митохондриальной и пластидной ДНК при взаимодействии со специфическими белками образуют нуклеоиды. Виды РНК и их строение В клетках организмов содержатся три вида РНК: рибосомная (рРНК), информационная или матричная (mРНК) и транспортная (тРНК). Основная функция РНК – перенос генетической информации от ДНК на белки. Первые сведения об участии РНК в синтезе белков были получены в 1941-42 г.г. Однако более полное представление о строении и функциях каждого из трёх видов РНК было достигнуто в 1956-1961 г.г., когда были открыты рибосомы и рибосомная РНК, а также выяснена биологическая роль информационной и транспортной РНК. Рибосомная РНК локализована в составе рибосом цитоплазмы и внутриклеточных органелл – митохондрий и хлоропластов. Однако её синтез происходит в ядре. У высших организмов местом синтеза большей части рРНК является ядрышко. Гены рРНК в хромосомах клеток представлены многими повторяющимися копиями. Полинуклеотидные последовательности рРНК содержат информацию о структуре рибосомных белков, многие из которых после их синтеза образуют с рРНК комплексы и химические соединения в виде нуклеопротеидов. В количественном отношении рРНК преобладает в клетке над другими видами РНК, на её долю приходится до 90% от общего содержания РНК в клетке. В клетках прокариот каждая рибосома содержит три молекулы рРНК, различающиеся по составу нуклеотидов и молекулярным массам. Большая по размерам субъединица рибосомы прокариот ( 50S-субъ-единица) содержит 23S-РНК, состоящую из 3300 нуклеотидных остатков, и 5S-РНК, включающую 120 нуклеотидных единиц. В малой субъединице находится 16S-РНК, в полинуклеотидной цепи которой насчитывается 1700 нуклеотидных остатков. В цитоплазматических рибосомах высших организмов содержатся четыре вида РНК: 28S-РНК, имеющие в своём составе 5000 нуклеотидных остатков (н.о.); 18S-РНК (2000 н.о.), 5,8S-РНК (150 н.о.) и 5S-РНК (120 н.о.). Гены, кодирующие структуру 28S-РНК, 18S-РНК и 5,8S-РНК локализованы в ядрышке, а генетическая информация о структуре 5S-РНК содержится в других участках хромосом вне ядрышка. В большой субъеденице цитоплазмотических рибосом высших организмов (60S-субъединица) локализованы 28S-РНК, 5,8S-РНК и 5S-РНК, в малой субъединице (40S-субъединица) – 18S-РНК. В хлоропластных рибосомах содержатся такие же по размерам молекулы рРНК, как и клетках прокариот. Митохондриальные рРНК по размерам и структуре также находятся ближе к бактериальным. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||