Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
 Скачать 7.39 Mb. Скачать 7.39 Mb.
|

|
Глава 5 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ Центральная нервная система координирует деятельность всех органов и систем, обеспечивает эффективное приспособление организма к изменениям окружающей среды, формирует целенаправленное поведение. Эти сложнейшие и жизненно важные задачи эешаются с помощью нервных клеток (нейронов), специализированных на восприятии, обработке, хранении и передаче информации и объединенных в специфически организованные нейронные цепи и центры, составляющие различные функциональные системы мозга. Объединение нервных клеток осуществляется с помощью синаптических соединений, важнейшей функцией которых является обеспечение перехода электрических сигналов г одного нейрона на другой. Число нервных элементов, будучи очень ограниченным у примитивных организмов, в процессе эволюционного развития нервной системы достигает многих миллиардов у приматов и человека. При этом количество синаптических контактов между нейронами приближается к астрономической цифре;—1015— 1016. Сложность организации ЦНС проявляется также в том, что структура и функция нейронов различных отделов мозга значительно варьируют. Тем не менее результаты исследования различных отделов мозга или клеток нервной системы животных, стоящих на разных уровнях эволюционного развития, позволяют выделить ряд общих закономерностей, определяющих течение основных нервных процессов: возбуждения и торможения в нейронах и синапсах ЦНС. Необходимым условием анализа деятельности мозга является выделение общих фундаментальных принципов, лежащих в основе функционирования нейронов и синапсов. НЕЙРОННАЯ ТЕОРИЯ В основе современного представления о структуре и функции ЦНС лежит нейронная ■теория, которая представляет собой частный случай клеточной теории. Однако, если клеточная теория была сформулирована еще в первой половине XIX столетия, то нейронная теория, рассматривающая мозг как результат функционального объединения отдельных клеточных элементов ■— нейронов, получила признание только на рубеже нынешнего века. До этого существовала тенденция рассматривать ЦНС как непрерывный синцитий, все элементы которого соединены прямой цитоплазматической связью. Большую роль в признании нейронной теории сыграли исследования испанского нейрогистолога Р. Кахала и английского физиолога Ч. Шеррингтона. Окончательные доказательства полной структурной обособленности нервных клеток были получены с помощью электронного микроскопа, высокая разрешающая способность которого позволила установить, что каждая нервная клетка на всем своем протяжении окружена пограничной мембраной и что между мембранами разных нейронов имеются свободные пространства. Нервная система построена из двух типовклеток: нервных и глиальных, причем число последних в 8—9 раз превышает число нервных. Однако именно нейроны обеспечивают все многообразие процессов, связанных с передачей и обработкой информации. Основные структурно-функциональные элементы нервной клетки. В каждой нервной клетке можно выделить четыре основных элемента (рис. 59): тело, или сому, дендриты, аксон и пре- синаптинеское окончание аксона. Каждый из этих элементов выполняет определенную функцию. Тело нейрона содержит различные внутриклеточные органеллы, необходимые для обеспечения жизнедеятельности всей клетки: ядро, рибосомы, эндо- плазматический ретикулум, пластинчатый комплекс (аппарат Гольджи), митохондрии. Здесь происходит основной синтез макромолекул, которые затем могут транспортироваться в дендриты и аксон. Мембрана тела большинства нейронов покрыта синапсами и, таким образом, играет важную роль в восприятии и интеграции сигналов, поступающих от, других нейронов. От тела клетки берут начало дендриты и аксон. В большинстве случаев дендриты сильно разветвляются. Вследствие этого их суммарная поверхность значительно превосходит поверхность тела клетки. Это создает условия для размещения на дендритах большого числа синапсов. Таким образом, именно дендритам принадлежит ведущая роль в восприятии нейроном информации. Мембрана дендритов, как и мембрана тела нейронов, содержит значительное число белковых молекул, выполняющих функцию химических рецепторов, обладающих специфической чувствительностью к определенным химическим веществам. Эти вещества участвуют в передаче сигналов с клетки на клетки и являются медиаторами синаптического возбуждения и торможения. Основной функцией аксона является проведение нервного импульса — потенциала действия. 1 Способность потенциала действия распространяться без ослабления, обеспечивает эффективное проведение сигнала по всей длине аксона, которая у некоторых нервных клеток достигает многих десятков сантиметров. Таким образом, основная задача аксона — проводить сигналы на большие .расстояния, связывая нервные клетки друг с другом и с. исполнительными органами. Окончание аксона специализировано на передаче сигнала на другие нейроны (или клетки исполнительных органов). Поэтому в нем содержатся специальные органеллы: синоптические пузырьки, или везикулы, содержащие химические медиаторы. Мембрана пресинаптических окончаний аксона в отличие от самого аксона снабжена специфическими рецепторами, способными реагировать на различные медиаторы или нёйромоду- ляторы. Благодаря указанному взаимодействию процесс выделения медиатора преси- наптическим окончанием может эффективно регулироваться другими нейронами. Кроме того, в отличие от остальной части аксона мембрана окончаний содержит значительное число кальциевых каналов, активация которых обеспечивает поступление внутрь окончания Са2+. 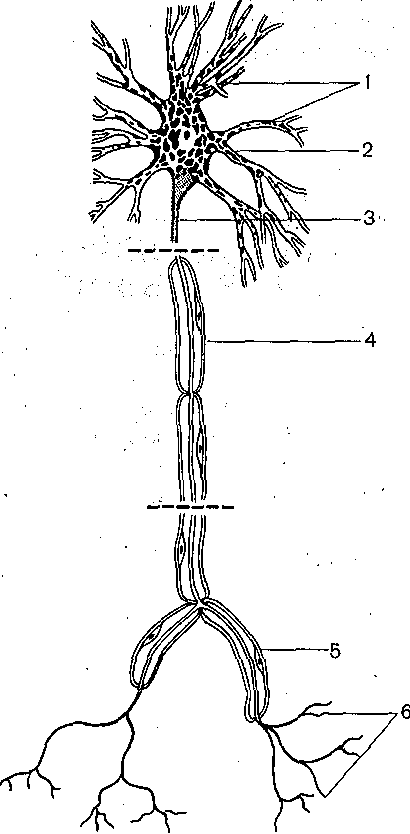 Рис. 59. Нервная клетка (схема). 1 — дендриты; 2 — тело клетки; 3 — эксон- пый холмик, 4 — аксон; 5.— коллатераль аксона; 6—пресинаптические окончания аксона. Типы нейронов. Количество отростков, выходящих из тела нервной клетки, может значительно варьировать. В соответствии с этим различают уни-, б и- и мультиполярные нейроны. Униполярные нейроны характерны главным образом для нервной системы .беспозвоночных. В нервной системе позвоночных имеются преимущественно би- и 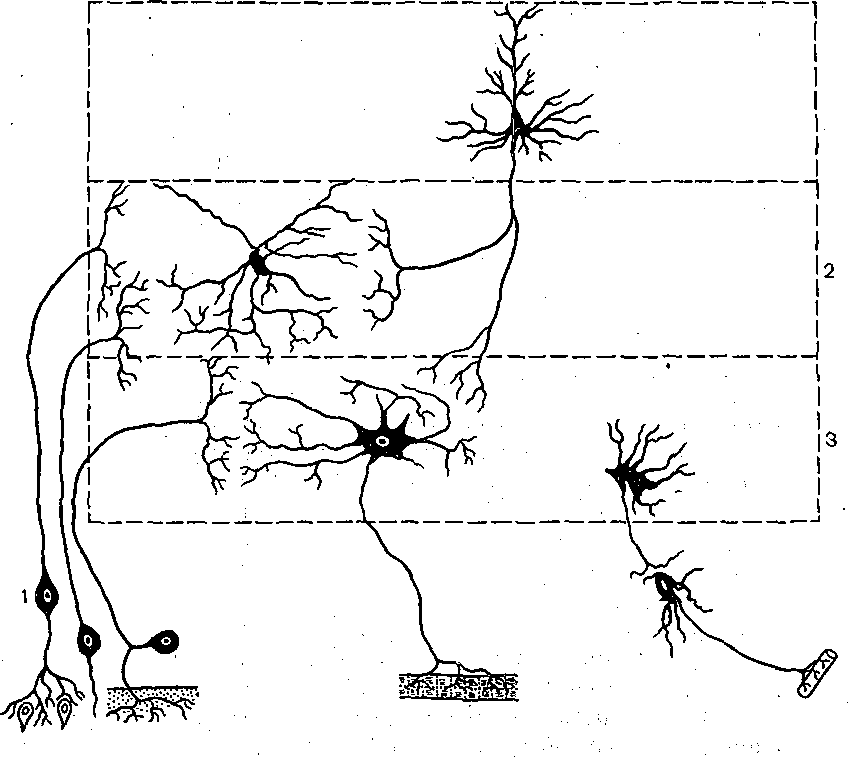 Рис. 60. Типы нервных клеток. 1 —сенсорные нейроны; 2— вставочные нейроны; 3 — эфферентные нейроны. мультиполярные нейроны. Последние особенно характерны для ЦНС. Тела биполярных нейронов обычно расположены на периферии, но их центральные отростки'вступают в ЦНС (рис. 60). Это так называемые первичные афферентные нейроны. Различают 3 основных типа нейронов: афферентные, вставочные и эфферентные. Первичные афферентные нейроны воспринимают сигналы, возникающие в рецепторных образованиях органов чувств, и проводят их в ЦНС. Вступая в пределы ЦНС, окончания отростков первичных афферентных нейронов устанавливают синаптические контакты со вставочными, а иногда и непосредственно с эфферентными нейронами." Вставочные нейроны локализуются, как правило, в пределах ЦНС. Они обеспечивают связь между различными афферентными и эфферентными нейронами. Аксоны эфферентных нейронов, например мотонейронов, выходят за пределы ЦНС и иннервируют волокна скелетной мускулатуры. Многие нейроны, которые можно отнести к эфферентным, передают сигналы не прямо на Периферию, а через посредство других нервных клеток. К таким эфферентным нейронам можно отнести нейроны различных отделов мозга, посылающие аксоны, идущие в составе длинных нисходящих трактов к спинному мозгу. Это пирамидные нейроны моторной зоны коры, руброспинальные, ретикулоспинальные и вестибуло- спинальные нейроны, импульсы от которых поступают к двигательным клеткам спин ал ь- ных моторных центров. Эфферентные нейроны вегетативной нервной системы расположены вне центральной нервной системы, в вегетативных ганглиях, находящихся на периферии. Их преганглионарные нейроны, локализованные в сером веществе мозгового ствола и спинного мозга, также относятся к эфферентным нейронам. Кабельные свойства мембраны. Все многообразие электрических сигналов, генерируемых, перерабатываемых и посылаемых любой нервной клеткой, может быть сведено всего лишь к двум типам: локальным (градуальным), потенциалам и потенциалам дей ствия (импульсным), Локальные потенциалы распространяются пассивно по кабельным структурам нейрона.,Поэтому по мере удаления от места своего возникновения они затухают и могут служить только для проведения сигналов на сравнительно небольшие расстояния, например от тела или дендритов нервной клетки к области начального сегмента аксона, где обычно, происходит процесс возникновения потенциала действия. Будучи значительно менее эффективным, чем потенциал действия, средством для передачи сигнала на расстояние, локальные потенциалы способны к суммации, и именно это свойство обеспечивает нейрону способность интегрировать все многочисленные поступающие к нему сигналы. Ввиду того что основным участком возникновения локальных потенциалов в нейронах ЦНС являются синапсы, которые расположены на мембране нейрона достаточно близко друг от друга, пространственное взаимодействие создаваемых синаптическими влияниями локальных процессов является достаточно эффективным.. . Потенциал действия, служащий для проведения сигналов на большие расстояния, благодаря наличию регенеративного механизма распространяется без ослабления. Здесь же следует подчеркнуть, что в целой нервной клетке благодаря ее сложному геометрическому строению и неодинаковым свойствам мембраны в различных участках процесс возникновения и распространения потенциала действия отличается рядом особенностей. Особенности возникновения и проведения потенциалов действия и локальных потенциалов. В большинстве нервных клеток порог возбудимости разных ее участков неодинаков. Он ниже всего в области аксонного холмика и начального сегмента аксона и выше в области сомы. Дендриты, как правило, имеют еще более высокий порог. Поэтому потенциал действия обычно возникает в области начального сегмента аксона и уже оттуда распространяется по аксону (ортодромно) и на тело клетки (антидромно). Если ввести в тело клетки микроэлектрод, позволяющий регистрировать потенциал действия, то можно видеть, что последний имеет характерную форму (рис. 61) , демонстрирующую наличие двух основных компонентов. Первый компонент обусловлен активацией зоны начального сегмента и аксонного холмика, второй — тела и деидритов нейрона. Задержка между, первым и вторым компонентами обусловлена тем, что более высокий порог возбудимости тела нейрона и значительное увеличение поверхности мембраны при переходе из аксонного холмика в тело нейрона затрудняют распространение потенциала действия на сомато-дендритическую мембрану. После окончания потенциала действия во многих нейронах ЦНС наблюдается длительная следовая гиперполяризация. Она особенно хорошо выражена в мотонейронах спинного мозга. 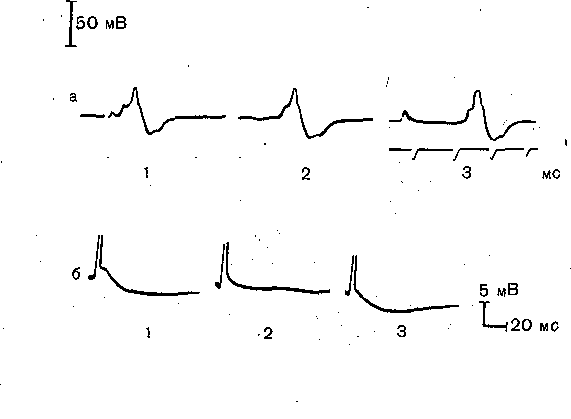 Рис. 61. Потенциал действия, регистрируемый микроэлектродом, введенным в тело мотонейрона. а — форма потенциала действия, вызываемого антидромно (1), синаптически (2) и прямым приложением тока через микроэлектрод (3); б — следовая гиперполяризация после потенциала действия мотонейрона котенка (1) и ее устранение после замены ионов кальция на ионы марганца (2) и восстановление в нормальном растворе (3)., Следовая гиперполяризация обусловлена тем, что соматическая мембрана в отличие от мембраны аксонов имеет значительное число кальциевых .каналов. Деполяризация мембраны, развивающаяся во время потенциала действия, активирует кальциевые каналы соматической мембраны (П. Г. Костюк). Входящие внутрь клетки ионы кальция в свою очередь активируют калиевую проводимость мембраны. "Активация калиевой проводимости выражается в развитии следовой гиперполяризации, наблюдаемой после окончания потенциала действия. Если заменить ионы кальция в окружающей нейроны среде на ионы марганца, для чего необходимо осуществлять изоляцию и перфузию участка мозга, следовая гиперполяризация обратимо блокируется {рис. 61, б). Следовая гиперполяризация играет важную роль в регуляции частоты потенциалов действия, генерируемых нервной клеткой. Способность нейрона отвечать ритмическими разрядами импульсов на длительную деполяризацию, создаваемую потоком импульсов, поступающих на его синапсы, представляет собой одну из важнейших характеристик его активности. В тех нейронах, где следовая гиперполяризация выражена значительно, частота импульсации не может быть очень высокой, так как ее верхние пределы ограничиваются, фактически рефрактерным периодом. Некоторые вставочные нейроны могут выдавать вспышки разрядов с частотой порядка 1000 в секунду. В мотонейронах спинного мозга длительность следовой гиперполяризации достигает 100—150 мс, что значительно увеличивает интервал между последующими потенциалами действия. Поэтому в обычных условиях частота ритмики мотонейронов не превышает 40—50 в секунду. Большинство двигательных актов осуществляется при еще более низкой частоте разрядов мотонейронов. Тонические мотонейроны имеют более длительную следовую гиперполяризацию и разряжаются с более редкой частотой, чем фазические мотонейроны, у которых следовая гиперполяризация короче. МЕХАНИЗМЫ СВЯЗИ МЕЖДУ НЕЙРОНАМИ Каждый многоклеточный организм, каждая ткань, состоящая из отдельных клеток, нуждается в механизмах, обеспечивающих межклеточные взаимодействия. Важное значение имеют процессы коммуникации клеток ЦНС. Главная задача их заключается в обработке и передаче информации, закодированной в виде электрических сигналов. Хотя межнейронные взаимодействия могли бы осуществляться различными путями (например, с помощью влияния электрических полей, генерируемых близко расположенными нервными элементами, изменением ионного состава среды вследствие перераспределения ионов в результате предшествующей активности, выделением в окружающую среду различных продуктов обмена и т. д.), в основе деятельности мозга лежат в основном механизмы, обеспечивающие передачуэлектрических сигналов с нейрона на нейрон через межклеточные соединения — синапсы, специализированные на передаче этих сигналов. Являясь главным механизмом связи между нейронами, синапсы во многом обеспечивают все многообразие функций мозга. Понятие синапс было введено в физиологию английским физиологом Ч. Шеррингто- ном (1897) для обозначения функционального контакта между нейронами. Следует отметить, однако, что еще в 60-х годах прошлого столетия И. М. Сеченов подчеркивал, что вне межклеточной связи нельзя объяснить способы происхождения даже самого элементарного нервного процесса. Чем сложнее устроена нервная система и чем больше число составляющих мозг нервных элементов, тем более важное значение имеют синаптические контакты. 1 Структура и функция синапсов. Различные синаптические контакты отличаются друг от друга механизмом действия, локализацией на поверхности клетки, функциональной направленностью (возбуждающие или тормозящие), способностью к модуляции в результате предшествующей активности. Однако при всем многообразии синапсов существуют определенные общие свойства их структуры и функции. Поэтому прежде чем рассматривать специфические особенности синапсов различных отделов ЦНС, необходимо описать общие принципы их функционирования. Синапс представляет собой сложное структурное образование, , в котором следует различать пресинаптическое звено или пресинапс (чаще всего это концевое разветвление аксона) и постсинаптическое звено или постсинапс (чаще всего участок мембраны тела или дендрита другого нейрона). Кроме наиболее распространенных типов межнейронных контактов — аксосоматических и аксодендритических, существуют также аксоаксонные, дендродендритические, сомато-дендритические и дендросоматинеские синапсы. Пресинаптическое окончание либо образует у постсинаптической клетки так называемые концевые бляшки, или бутоны, либо формирует по своему ходу многочисленные последовательные зоны контакта с различными участками постсинаптического нейрона (так называемые проходящие синапсы). Механизм передачи через синапс долгое время оставался невыясненным, хотя было очевидно, что передача сигналов в синаптической области резко отличается от процесса проведения потенциала действия по аксону. В начале XX в, была четко сформулирована альтернатива: синаптическая передача осуществляется или электрическим, или химическим . путем. Электрическая теория синаптической передачи в ЦНС пользовалась признанием до начала 50-х годов, хотя она и значительно сдала свои позиции после того, как химический механизм передачи был продемонстрирован в ряде периферических синапсов. Перфузия верхнего шейного симпатического ганглия (А. В. Кибяков), а также использование микроэлектродной техники для внутриклеточной регистрации синаптических потенциалов нейронов ЦНС (Экклс) позволили сделать вывод о химической природе передачи в межнейрональных синапсах спинного мозга. Эти факты послужили основанием для вывода об универсальности химического механизма передачи во всех синапсах ЦНС. Микроэлектродные исследования последних лет показали, однако, что в определенных межнейронных синапсах существует электрический механизм передачи. В настоящее время стало очевидным, что есть синапсы как с химическим, так и электрическим механизмом передачи. Более того, в некоторых синаптических структурах сочетанно функционируют и электрический и химический механизмы передачи (смешанные синапсы, или синапсы двоякого действия). Синапсы с электрическим механизмом передачи чаще встречаются у животных с более примитивной нервной системой, хотя они и обнаружены в мозге млекопитающих, включая приматов. Их число уменьшается в процессе эмбрионального развития. Синапсы с химическим механизмом передачи составляют большую часть синаптического аппарата ЦНС высших животных и человека. |