|
|
Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
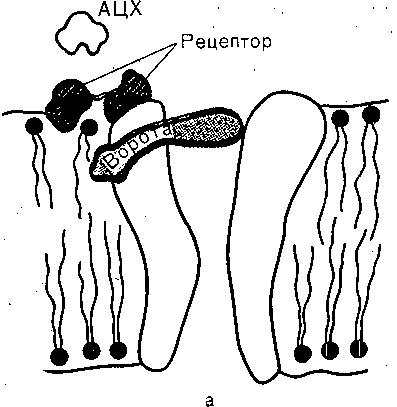
Механизмы возникновения ПКП и ПД в мышечном волокне. Хемовозбудимые каналы. ПКП обусловлен активацией ацетилхолином хемовозбудимых ионных каналов, шеющихся в постсинаптической мембране скелетного мышечного волокна. Так же как (лектровозбудимые, хемовозбудимые каналы образованы макромолекулами белка, пролизывающими липидный бислой мембраны. Функциональная структура хемовозбуди- лого канала схематически показана на рис. 53. Канал состоит из транспортной системы, юротного механизма и участка связывания — «холинорецептора», обладающего шсоким сродством к медиатору ацетилхолину. В отсутствие ацетилхолина канал закрыт, взаимодействие медиатора с рецептором приводит к активации канала. В открытом состоянии канал имеет проводимость порядка 30 пк Смс, что примерно В 4. раза превышает фоводимость одиночного электровозбудимого натриевого канала. Судя по величине
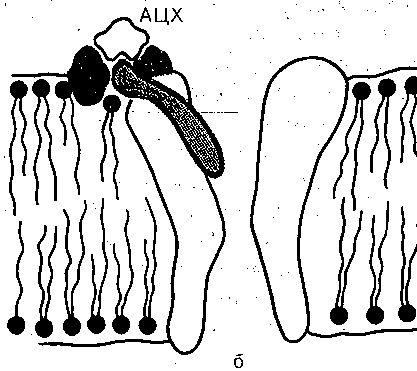
Рис-. 53. Схематическое изображение хемовозбудимого ионного канала, активизируемого ацетилхо лином. Канал образован макромолекулой белка, погруженной в липидный бислой мембраны. Воро та канала управляются хеморецептором (заштрихован). До взаимодействия молекулы ацетилхоли на (АЦХ) рецептором ворота закрыты (а). Они открываются при связывании АЦХ с рецептором (б'
наибольшего катиона, проникающего через рассматриваемые хемовозбудимые каналь размер их «пор» составляет примерно 0,65 нм, т. е. существенно превышает размер «пор в электровозбудимых натриевых и калиевых каналах. Соответственно ионная селектив ность хемовозбудимых каналов значительно ниже, они хорошо проницаемы и для ионо Na+ и для ионов Са2+, К+. Эти ионы движутся через открытые каналы по концентрацией ному и электрическому градиентам, и, поскольку внутреннее содержимое мышечног волокна заряжено электроотрицательно по отношению к наружному раствору, пото положительно заряженных' ионов Na+ внутрь клетки превышает противоположи направленный поток ионов К+. В;результате мембрана деполяризуется. Эта деполяр^ зация постсинаптической мембраны имеет, однако, нерегенеративный характер, пс скольку хемовозбудимые каналы не обладают электровозбудимостью: порция ацети-г холина, поступившая к постсинаптической мембране, активирует определенное числ! хемовозбудимых каналов. Это вызывает деполяризацию мембраны, но такая деполяр^ зация не способствует дальнейшему увеличению числа активируемых каналов. Поэтом значение ПКП зависит от концентрации ацетилхолина, действующего на мембран} чем больше эта концентрация, тем выше до определенного предела ПКП. Таким образоь ПКП в отличие от потенциала действия градуален. В этом отношении он сходен с локал! ным ответом, хотя имеет иной механизм.
Между деполяризованной ацетилхолином постсинаптической мембраной и сосед ними с ней участками электровозбудимой мембраны скелетного мышечного волоки возникают местные токи, вызывающие генерацию потенциала действия, распространяю щегося по всему мышечному волокну. Условием возникновения этого потенциала деЈ ствия является критическая деполяризация электровозбудимой мембраны, происходя щая при достижении ПКП пороговой величины.
Процесс передачи возбуждения с нервного волокна на скелетное мышечное може
быть схематически изображен в виде следующей цепи явлений: нервный импульс
поступление ионов Са2+ внутрь нервного окончания освобождение изокончани
ацетилхолина взаимодействие ацетилхолина с холинорецептором активг
ция хемовозбудимых каналов постсинаптической мембраны в результате взаимодействи
медиатора с холинорецепторами >- возникновение потенциала концевой плг
стинки критическая деполяризация околосинаптической электровозбудимо
мембраны -»- генерация потенциала действия.
Миниатюрные постсинаптические потенциалы
Ацетилхолин еекретируется двигательными нервными окончаниями не только при возбуждении, но и в покое. Различие состоит лишь в том, что в покое выделяются малые порции — «кванты» — ацетилхолина, а под влиянием нервного импульса в синаптИче- скую щель одновременно выбрасывается- значительное количество таких «квантов». «Квант» представляет собой «пакет» молекул медиатора в единичном пузырьке нервного окончания, изливающем свое содержимое в синаптическую щель. В концевой пластинке различных животных в каждом «кванте» содержится до. 2000 молекул ацетилхолина. Выделение отдельных квантов в синаптическую щель в состоянии покоя вызывает кратковременную слабую деполяризацию постсинаптической мембраны мышечнбго волокна.. Такая деполяризация получила название миниатюрного потенциала, поскольку она по своей амплитуде (0,5 мВ) в 50—80 раз меньше ПКП, вызываемого одиночным нервным импульсом. Миниатюрные потенциалы возникают обычно с частотой примерно один в секунду, они зарегистрированы не только в нервно-мышечных соединениях, но и в синапсах нервных клеток ЦНС.
Влияние кураре на нервно-мышечное соединение
Существует ряд веществ, также обладающих сродством к холинорецептору, но образующих с ним более прочную связь, чем ацетилхолин. К числу таких веществ относятся кураре и некоторые другие соединения (д-тубокурарин, диплацин, флакседил). После их воздействия на мышцу холинорецептор оказывается заблокированным и ни нервный импульс, ни искусственно введенный'ацетилхолин не способны вызвать возбуждение мышечного волокна.
Многие годы изучение действия кураре на нервно-мышечную передачу представляло только теоретический интерес, и физиологи были очень далеки от мысли, Что этот препарат может когда- 1ибо найти применение в медицинской практике. Однако в связи с развитием хирургии возникла 1еобходимость изыскания средств, которые позволили бы проводить оперативные вмешательства i условиях выключения естественного дыхания. И здесь кураре и его производные оказали большую юмощь. В настоящее время многие полостные операции проводят в условиях искусственного хыхания на фоне нервно-мышечной блокады препаратами, действующими подобно кураре.
Исключительно прочную связь с холинорецептором образует токсин из яда змеи —■ а-бунга- ютоксин. Этот токсин, снабженный радиоактивной меткой, позволил выделить холинорецептор из лембрэны. Химический анализ холинорецептора показал, что холинорецептор является липопро- 'еидом с молекулярной массой около 300 000.
Холинэстераза и ее роль
в процессах нервно-мышечной передачи
Установлено, что в области нервно-мышечного соединения в больших концентра- ,иях присутствует фермент холинэстераза, способная быстро расщеплять ацетилхолин, ыделяющиЙся в нервном окончании. Значение этого процесса становится ясным, если честь, что в естественных условиях к мышце поступают быстро следующие друг за дру- ом нервные импульсы и постсинаптическая мембрана, деполяризованная предшествующей порцией ацетилхолина, становится малочувствительной к действию следующей орции. Чтобы идущие друг за другом нервные импульсы могли осуществлять нормаль- ое возбуждающее действие, необходимо к моменту прихода каждого из них «убрать» редшествующую порцию медиатора. Эту. функцию и выполняет холинэстераза. ХоЛин, свобождаюйцийся при расщеплении молекул ацетилхолина, переносится обратно в ервное окончание специальной транспортной системой, существующей в пресинапти- еской мембране.
Существует ряд. фармакологических агентов, обладающих способностью резко угне- ать активность холинэстеразы. Их называют ингибиторами. К числу таких веществ
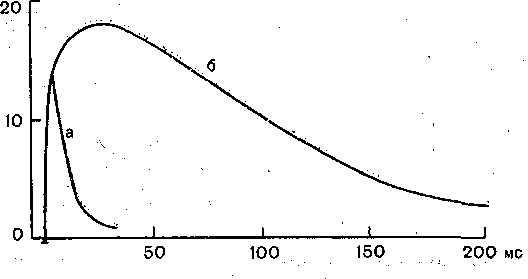
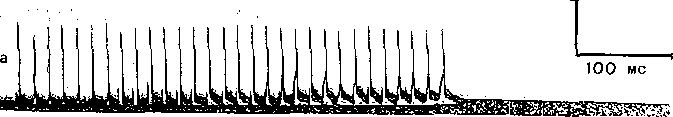
Рис. 54; Влияние ингибитора холинэстеразы (нео,- Рис. 55. Оптимум (Ор) и пессимум (Ps
стигмина) на длительность постсинаптического в нервно-.мышечном аппарате,
потенциала (потенциала концевой пластинки) Цифры обозначают частоту раздражения, одиночного мышечного волокна.
а — до применения,неостигмина; б— после применения неостигмина.
относятся эзерин, простигмин, галантамин. Если на нервно-мышечное соединени действует какоеглибо из этих веществ, постсинаптический потенциал увеличиваете по своей амплитуде и резко растягивается по времени.
Иллюстрацией этого является приведенная на рис. 54 запись ПКП, зарегистрирс ванного в нервно-мышечном соединении лягушки до (а) и после (б) воздействия н мышцу вещества, угнетающего активность холинэстеразы.
При действии ингибитора холинэстеразы ритмическое раздражение нерва вызывае выраженную суммацию ПКП, что ведет к стойкой деполяризации постсинаптическо мембраны и блоку проведения импульсов с нервного волокна на мышечное. При это стойкая деполяризация постсинаптической мембраны, приводит соседние участк мышечного волокна в состояние угнетения, обусловленное инактивацией натриево и стойким повышением калиевой проводимости мембраны (состояние «катодическо депрессии»).
Следует отметить, что и в отсутствие ингибиторов холинэстеразы при условии очек частого ' раздражения нерва постсинаптические потенциалы / (ПКП), вызываемь каждым нервным импульсом, суммируются, поскольку в межимпульсный интерва холинэстераза не успевает полностью расщепить выделяющийся в нервном окончанн ацетилхолин. В результате суммации потенциалов постсинаптическая мембрана в< более и более деполяризуется.
ПЕССИМАЛЬНОЕ ТОРМОЖЕНИЕ
Деполяризация постсинаптической мембраны при очень частом следовании дру за другом нервных импульсов лежит в основе открытого Н. Е, Введенским пессимальног торможения. Это явление часто называют торможением Введенского. Сущность ег состоит в следующем. Величина тетанического сокращения скелетной мышцы в отве на ритмические раздражения нерва возрастает с увеличением частоты стимуляции. Пр некоторой оптимальной частоте раздражения тетанус достигает наибольшей величинь Если продолжать увеличивать частоту стимуляции нерва, то тетаническое сокращени мышцы начинает резко ослабевать и при некоторой большой пессимальной часто! раздражения нерва мышца, несмотря на продолжающееся раздражение, почти noj ностью расслабляется. Уменьшение частоты стимуляции тотчас приводит к восстановл< нию высокого уровня тетанического сокращения (рис. 55).
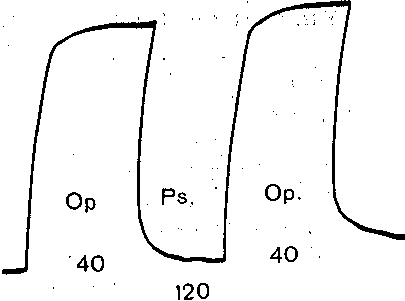
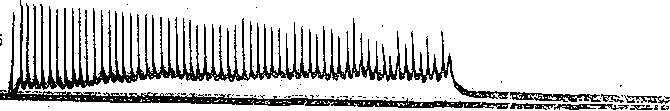
В нервно-мышечном препарате лягушки торможение Введенского возникает пр частоте раздражения, близкой к 100 стимулам в секунду, т. е. при значительно меньши
Рис. 56. Суммация постсинапти-^ ческих потенциалов и стойкая деполяризация постсинаптиче: ской мембраны мышечного волокна при пессимальном раздражении.
50 (и В
дшшюшжшлаладш
Внутриклеточные отведения от си- наптической области. Частота раздражения: а — 65 в секунду; б — 125 в секунду; в — 150 в секунду; г — 180 в секунду. Обозначения кривых сверху вниз: потенциалы мышечного волокна, исходный уровень потенциала покоя; потенциалы действия нерва {по Н. М. Шамариной).
 :астотах, чем те, которые может воспроизводить нерв (порядка 500) или мышца (порядка 200) На рис. 56 приведена запись изменения мембранного потенциала мышечного во- юкна в области концевой пластинки при раздражении церва ритмическими импульсами >птимальной и пессимальной частот. При редком (оптимальном) ритме каждый [мпульс попадает в фазу убывания постсинаптического потенциала, вызванного преды- 1ущим импульсом, и в мышечном волокне возникают полноценные потенциалы действия. 1ри частом раздражении постсинаптические потенциалы суммируются, что приводит : стойкой деполяризации постсинаптической мембраны и развитию блока проведения. ■1а важную роль ацетилхолина в развитии пессимума Введенского указывает тот факт, [то яды, инактивирующие холинэстеразу и тем самым способствующие накоплению ! области синапса ацетилхолина, способствуют возникновению пессимума. Следует подчеркнуть, что рассмотренный механизм блокирования нервно-мышеч- ого соединения при частом ритме раздражения нерва не является единственным. В тех лучаях, когда частота стимулов очень высока, проведение возбуждения с нерва на гышцу может быть блокировано еще на пути к синапсу, в тонких пресинаптических 'азветвлениях нервных волокон — пресинаптических терминалях, обладающих более изкой лабильностью, чем толстые нервные волокна. Нарушение проведения в этих онких нервных волокнах ведет к прекращению поступления нервных импульсов к нерв- :ому окончанию и тем самым к прекращению выделения ацетилхолина. В данном случае место стойкой деполяризации постсинаптической мембраны обнаруживается значи- ельное ослабление или даже полное выпадение постсинаптических потенциалов при еизменном уровне потенциала покоя мышечного волокна. Нарушение нервно-мышечной передачи при утомленииПри длительном раздражении нерва нарушение нервно-мышечной передачи развивается адолго до того, как мышца, а тем более нерв в силу утомления утрачивают способность к проведе- ию возбуждения. Объясняют это тем, что в нервных окончаниях при длительном раздражении уменьшается запас «заготовленного» медиатора. Поэтому порции ацетилхолина, выделяющиес! в синапсах в ответ на каждый импульс, уменьшаются и соответственно снижаются до подпороговы: величин постсинаптические потенциалы.. Наряду с этим при длительном раздражении нерва по; влиянием продуктов обмена в мышце происходит постепенное понижение чувствительности пост синаптической мембраны к ацетилхолину. В результате величина потенциалов концевой пластиню уменьшается. Когда амплитуда ПКП падает ниже некоторого критического'уровня, возникновени! потенциалов действия в мышечном волокне прекращается. По этим причинам синапсы быстре' утомляются, чем нервные волокна и мышцы. ТРОФИЧЕСКАЯ ФУНКЦИЯ ДВИГАТЕЛЬНЫХ НЕРВНЫХ волокон И ИХ ОКОНЧАНИЙ Наряд/ с функцией передачи импульсов, вызывающих мышечные сокращения нервные волокна и их окончания оказывают также трофическое воздействие на мышцу т. е. участвуют в регуляции ее обмена веществ. Хорошо известно, что денервация мышцы развивающаяся при дегенерации двигательного нерва, приводит к атрофии мышечньп волокон, которая проявляется в том, что вначале уменьшается количество саркоплазмы а затем и диаметр мышечных волокон; позднее происходит разрушение миофибрилл Специальные исследования показали, что эта атрофия не является результатом лиил бездеятельности мышцы, потерявшей двигательную активность. Бездеятельност] мышцы может быть вызвана и путем тендотомии, т. е. перерезки сухожилия. Однако если сравнить мышцу после тендотомии и после денервации, можно убедиться, что е последнем случае в мышце развиваются качественно иные изменения ее свойств, ш обнаруживающиеся щри тендотомии. Наиболее ярко это проявляется в изменения) чувствительности мышцы к ацетилхолину. В нормальной и тендотомированной мышц( к ацетилхолину чувствительна только постсинаптическая мембрана, в которой сосре доточены хемовозбудимые ионные каналы, снабженные холинорецепторами. Денервации приводит к тому, что такие же каналы появляются и во внесинаптических областям мышечного волокна. В результате чувствительность денервированной мышцы к ацетил холину резко возрастает. Указанная гиПерчувствительность к ацетилхолину не форми руется, если при помощи определенных химических реагентов затормозить белковьп синтез в мышечных волокнах. Реиннервация мышцы вследствие регенерации нервны) волокон приводит к исчезновению холинорецептивных каналов области внепостсинапти ческой мембраны. Эти данные свидетельствуют о том, что нервные волокна регулируют синтез белков, образующих хемовозбудимые холинорецепторны'е. каналы. В денервированной мышце резко падает также активность ряда ферментов, в частности АТФ-азы, играющей важную роль в процессе освобождения' энергии, заключенной в фосфатных связях АТФ. В то же время при денервации значительно усилены процессы распада белков. Это приводит к характерному для атрофии постепенному уменьшению массы мышечной ткани^ Все дегенеративные изменения в денервированной мышце начинаются'тем раньше, чем на меньшем расстоянии от мышцы перерезают двигательный нерв; Это позволяет предположить, что определенные вещества («трофические агенты»), вырабатываемые в нервных клетках, продвигаются по нервным волокнам от проксимальных участков к дистальным и выделяются нервными окончаниями. Чем больший отрезок нерва остается соединенным с мышцей, тем дольше она получает важные для ее обмена вещества. Перемещение этих веществ осуществляется благодаря движению нейроплазмы, скорость которого 1—2 мм/ч. Важную роль в осуществлении трофических влияний нерва играет ацетилхолин, секретируе- мый нервными окончаниями как в покое, так особенно при возбуждении. Имеются основания считать, что ацетилхолин и продукты его расщепления холинэстеразой — холин и уксусная кислота — участвуют в обмене веществ мышцы, оказывая активирующее влияние на определенные ферментные системы. Так, при введении ацетилхолина в денервированную мышцу кролика резко увеличивается распад аденозинтрифосфата, креатинфосфата и гликогена во время тетануса, вызванного прямым электрическим раздражением этой мышцы. Из нервных окончаний выделяются вещества, которые оказывают специфическое влияние на синтез белков мышечного волокна. Об этом свидетельствуют опыты с перекрестным сшиванием двигательных нервов, иннервирующих быстрые и медленные скелетные мышцы. При таком сшивании периферические отрезки нервов и их окончания в мышце дегенерируют, а по их путям в мышцу прорастают новые волокна из центральных отрезков Нервов. Вскоре после того, как эти волокна образуют двигательные окончания, происходит отчетливая перестройка функциональных свойств мышц. Мышцы, которые ранее были быстрыми, теперь становятся медленными, а те, которые были медленными, становятся быстрыми. При такой перестройке изменяется активность АТФ-азы их сократительного белка миозина: в бывших быстрых мышцах она резко падает, а в медленных возрастает. Соответственно в первых скорость распада АТФ увеличивается, а во вторых — уменьшается. Изменяются также свойства ионных каналов клеточной мембраны. Трофическое влияние на скелетную мышцу оказывают и волокна симпатической нервной системы, окончания которых высвобождают норадреналин. ОСОБЕННОСТИ НЕРВНО-МЫШЕЧНОЙ ПЕРЕДАЧИ ВОЗБУЖДЕНИЯ В ГЛАДКИХ МЫШЦАХМеханизм передачи возбуждения с двигательного нервного волокна на волокна, гладкой мышцы в принципе сходен с механизмом нервно-мышечной передачи в скелетной мускулатуре. Различия касаются лишь химической природы медиатора и особенностей суммации постсинаптических потенциалов. Во всех скелетных мышцах возбуждающим медиатором является ацетилхолин. В гладких мышцах передача возбуждения в нервных окончаниях осуществляется при помощи разных медиаторов. Тйк, для гладких мышц желудочно-кишечного тракта возбуждающим медиатором является ацетилхолин, а для гладких-мышц кровеносных сосудов — норадреналин. Порция медиатора, высвобождаемая нервным окончанием в ответ на одиночный нервный импульс, в большинстве случаев оказывается недостаточной для критической деполяризации мембраны гладкомышечной клетки. Критическая деполяризация происходит только при поступлении к нервному окончанию нескольких следующих друг за другом импульсов. Тогда одиночные возбуждающие постсинаптические потенциалы суммируются (рис. 57) и в момент, когда их сумма достигает пороговой величины, возникает потенциал действия. В скелетном мышечном волокне частота следования потенциалов действия соответствует частоте ритмического раздражения двигательного нерва. В отличие от этого в гладких мышцах такое соответствие нарушается уже при частотах 7—15 имп/с. Если же частота стимуляции превышает 50 имп/с, возникает торможение типа пессимального. Тормозные синапсы в гладких мышцах. Раздражение некоторых нервных волокон, иннервирующих гладкие мышцы, может вызывать их торможение, а не возбуждение. Нервные импульсы, приходящие в определенные- нервные окончания, высвобождают тормозной медиатор. Воздействуя на постсинаптическую мембрану, тормозной медиатор взаимодействует с хемовозбудимыми каналами, обладающими преимущественной проницаемостью для ионов К +. Выходящий поток калия через эти каналы вызывает гиперполяризацию постсинаптической мембраны, проявляющуюся в форме «тормозного постсинаптического потенциала», подобного тому, который наблюдается в тормозных синапсах нейронов в ЦНС. При ритмическом раздражении тормозных нервных волокон тормозные постсинаптические потенциалы суммируются друг с другом, причем эта суммация оказывается наиболее эффективной в диапазоне частот 5—25 имп/с (рис. 58). Если раздражение тормозящего нерва несколько предшествует стимуляции активирующего нерва, то возбуждающий постсинаптический потенциал, вызываемый по- 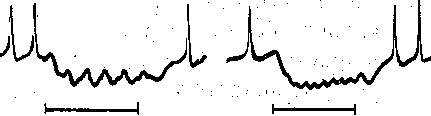 50 мВ 1— 1— к 1 Секунды .
Рис. 58. Тормозные постсинаптические потенциалы в гладкомышечном волокне кишки морской свинки, а — тормозный потенциал, вызванный одиночным раздражением интрамурального тормозного нерва; б и в — суммация тормозных потенциалов при ритмическом раздражении этого нерва. Частота раздражений 2 в секунду (б) и 4 в секунду (в). Горизонтальная линия— период раздражения.
следннм, ослабляется и может оказаться недостаточным для критической деполяризация мембраны. Раздражение тормозного нерва на фоне спонтанной активности мышцы угнетает генерацию потенциалов действия и, следовательно, приводит к прекращению ее сокращений. Роль тормозного медиатора в гладких мышцах, возбуждаемых ацетилхолином (например, кишечника, бронхов), исполняет норадреналин. Наоборот, в мышечных клетках сфинктера мочевого пузыря и некоторых других гладких мышцах, для которых возбуждающим медиатором является норадреналин, тормозным, медиатором служит ацетилхолин. Последний оказывает тормозящее действие и на клетки водителя ритма сердца. В скелетных мышцах нервно-мышечная передача, осуществляемая при помощи ацетилхолина, блокируется препаратами кураре, обладающими большим сродством к холинорецепторам. В гладких мышцах холинорецептор имеет иную химическую структуру, чем в скелетных, поэтому она блокируется не препаратами кураре, а атропином. В тех гладких мышцах, в которых медиатором служит норадреналин', хемовозбудимые каналы снабжены адренорецепторамй. Различают два основных вида адренорецеп- торов: а-адренорецепторы и $-адренорецепторы, которые блокируются различными химическими соединениями — адреноблокаторами. ЗАКЛЮЧЕНИЕК возбудимым тканям кроме нервной и мышечной относится и железистая ткань, но механизмы возбуждения клеток желез внешней секреции несколько отличны от таковых у нервных и мышечных. 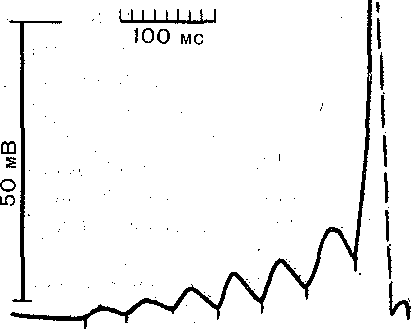
Рис. 57. Суммация постсинаптических потенциалов в гладкомышечных волокнах морской свинки.
Как показали микроэлектродные исследования мембрана секреторных клеток в состоянии покоя является поляризованной, причем наружная поверхность ее заряжена положительно, а внутренняя — отрицательно. Разность потенциалов составляет 30-— 40 мв. При стимуляции секреторных нервов, иннервирующих железу, возникает не деполяризация, а гиперполяризация мембраны и разность потенциалов достигает 50—60 мв. Предполагают, что это происходит вследствие нагнетания С1 и других отрицательных ионов в клетку. Под влиянием электростатических сил в клетку вслед за этим начинают поступать положительные ионы, что приводит к повышению осмотического давления, поступлению в клетку воды, увеличению гидростатического давления и набуханию клетки. В результате возникает выброс секрета из клетки в просвет жеЛезы. . Выброс секрета может стимулироваться не только нервными, но и химическими (гуморальными) влияниями. Здесь, как и везде в организме, регуляция функций осуществляется двумя способами — нервным и гуморальным. Нервный импульс представляет собой наиболее быстрый способ передачи информации в организме. Поэтому в процессе эволюции в тех случаях, когда была необходима большая скорость реакций, когда от быстроты ответных реакций зависело само существование организма, этот способ передачи сигналов стал основным. В области нервных окончаний — в синаптических щелях нервный импульс, как правило, вызывает выделение медиатора и, таким образом, взаимодействие между клетками остается по существу химическим. При этом вместо медленного распространения химического вещества с током жидкости (с движущейся кровью, лимфой, тканевой жидкостью и т. д.) в нервной системе с большой скоростью распространяется сигнал к выделению биологически активного вещества (медиатора) в области нервных окончаний (на месте). Все это резко повысило быстроту ответных реакций организма, сохранив по существу принцип химического взаимодействия между клетками. Вместе с тем в ряде случаев, когда при клеточном взаимодействии необходима еще более быстрая и притом всегда однозначная реакция, межклеточная передача сигнала обеспечивается прямым электрическим взаимодействием клеток. Такой тип связи наблюдается, например, при взаимодействии клеток миокарда, а также некоторых электрических синапсов ЦНС, получивших название э^апсов. Межклеточные связи сводятся не только к электрическим взаимодействиям или влияниям медиаторов. Химическая взаимосвязь между клетками является более сложной. Клетки органов и тканей вырабатывают, ряд специфических химических веществ, действующих на другие клетки и вызывающих не только включение и выключение (или усиление или ослабление) функции, но и изменение интенсивности обмена веществ и процессов синтеза клеткой специфических белков. Механизмы всех этих рефлекторных влияний и межклеточных взаимодействий подробно рассмотрены во втором разделе учебника. Раз дел II МЕХАНИЗМЫ РЕГУЛЯЦИИ ФИЗИОЛОГИЧЕСКИХ ПРОЦЕССОВ ВВЕДЕНИЕЧеловеческий организм представляет собой систему (грубее говоря — машину) — единственную по высочайшему саморегулированию. С этой точки зрения, метод изучения системы человека тот же, как и всякой другой системы: разложение на части, изучение значения каждой части, связи частей, соотношений с окружающей средой, и, в конце концов, понимание на основе всего этого ее общей работы и управление ею (И. П. Павлов) . В этих словах выражено содержание понятия системного подхода.Системный подход представляет собой Методологию научного познания, в основе которого лежит рассмотрение объектов как систем. Этот подход ориентирует исследователя на раскрытие целостности объекта, на выявление многообразных типов связей в нек и на создание общего представления о системе. Объекты высокой степени сложности к которым относится организм человека, представляют собой многоуровневую органи зацию, в которой системы более высокого уровня и сложности включают в себя системь более низкого уровня, образуя иерархию подсистем. Связи элементов в системе любогс уровня осуществляются, путем передачи информации. В организме, животных и человеке информация закодирована в определенной структуре биологических молекул, а также в определенном «рисунке» нервных импульсов (частота, набор в пачки, интервалы межд} пачками, определенное соотношение во времени импульсов и их пачек в различны) нервных волокнах и т. д.). С помощью передачи этой информации осуществляются процессы регуляции, т. е, управления физиологическими функциями, деятельностью клеток, тканей, органов, систем, поведением организма, осуществление взаимодействия организма и окружающей среды. , Главным регуляторным (управляющим) механизмом в организме высших животных и человека является нервная система. Основной механизм ее деятельности — рефлекс. Рефлексом (от лат, reflecto — отражение) называют любую ответную реакцию организма, осуществляющуюся с участием центральной нервной системы. Морфологической основой таких реакций является рефлекторная дуга, включающая 5 звеньев: 1) рецептор — специализированный прибор, воспринимающий определенный вид воздействий внешней или внутренней среды; 2) афферентный (чувствительный) нейрон (или нейроны), проводящий сигнал, возникающий в рецепторе, в нервный центр; 3) вставочный нейрон (или нейроны), представляющий собой центральную часть рефлекторной дуги, (или нервный центр) указанного рефлекса; 4) эфферентный (двигательный) нейрон, по аксону которого сигнал доходит до эффектора; 5) эффектор — поперечнополосатая или гладкая мышца либо железа, осуществляющие соответствующую деятельность. Любой эффектор, таким образом, связан элементами рефлекторной дуги с соответствующим рецептором и запускается в действие при раздражении данного рецептора. Ответная реакция организма возникает вследствие распространения по рефлекторной дуге возбуждения (сигнала), появляющегося при раздражении рецептора. Понятие о рефлексе было введено в середине XVI века великим французским ученым Рене Декартом. Введение этого понятия сыграло важнейшую роль в развитии физиологии, позволило объяснить причину ответных реакций организма, изучить их механизм и показать, что в основе таких реакций лежит принцип детерминизма (т. е. всеобщий как для неживой, так и для живой природы принцип причинно-следственных отношений). Тем самым был сделан важный шаг в развитии материалистических представлений' о. механизме реакций организма.- Со времен Декарта подобные реакции считались машинообразными, обеспечивающими автоматизированный ответ организма на раздражение рецептора. Однако подобные автоматизированные реакции имеют место лишь при возникновении элементарных простых рефлексов, которые могут осуществляться с участием ограниченных звеньев ЦНС. Как правило, рефлекторные реакции организма являются гораздо более сложными и происходят при участии многих звеньев (этажей) ЦНС. Рефлексы при этом не сводятся к простым, однозначным ответным реакциям, а представляют собой звенья сложного процесса управления двигательными функциями (поведением) или деятельностью внутренних органов. Функциональная структура таких процессов управления (регуляции) намного сложнее, нежели структура отдельных машинообразных рефлекторных ответов. Процессам управления независимо от того, где бы они не осуществлялись в организме животного или человека, производственном процессе, социальном обществе и т. д., присущи некоторые общие черты и закономерности. Эти общие черты исследуются наукой, получившей название кибернетика. Кибернетика изучает общие черты и законы управления, осуществляемого на основе получения, хранения, передачи и переработки информации, независимо от физической природы объекта или системы, в которых осуществляются эти процессы. Кибернетическими системами могут быть автоматические регуляторы в технике, ЭВМ, организм человека и животных, биологическая популяция, человеческое общество. Изучение законов кибернетики, понимание их смысла весьма важно для познания сущности процессов регуляции физиологических функций, для моделирования (математического или экспериментального) этих функций, для автоматического контроля за осуществлением этих функций, для вмешательства в физиологические процессы с целью их нормализации в случаях расстройств и заболеваний. Изучение механизмов регуляции физиологических процессов раскрывает общность принципов кибернетики для всех указанных объектов, 1 единство принципов, автоматического регулирования в организме, в машине и производственном процессе. Известно, что сами процессы управления и автоматического регулирования были использованы в технике гораздо раньше, чем они были открыты в организме, и до того, как были сформулированы законы кибернетики. В машинах существуют «регуляторы, которые заменяют руку машиниста, приходя в целесообразную деятельность, как говорится сами собой, но в сущности под влиянием изменяющихся условий в ходе машины. Таков, например, предохранительный клапан в паровиках (паровых машинах) Уатта. По мере того, как напряжение пара в котле возрастает за известный предел, клапан сам собой увеличивает отверстие для выхода пара и наоборот. Таких приспособлений известно множество и все они носят название автоматических регуляторов. В животном теле, как в самодействующей машине, регуляторы, очевидно, могут быть только автоматическими, т. е. приводится в действие измененными условиями в состоянии или ходе машины (организма) и развивать деятельности, которыми эти неправильности устраняются» — писал И. М. Сеченов еще в 1897 году, предвосхищая положения кибернетики о механизмах саморегуляции в организме. Таким образом, И. М. Сеченовым был сформулирован принцип отрицательной обратной связи, лежащий в основе процессов автоматического регулирования в машине и живом организме. По этому принципу регулируются многие физиологические процессы. На значение этого факта впервые обратил внимание Клод Бернар (французский физиолог и патолог), обнаруживший значение постоянства внутренней среды для жизни-организма. На примере регуляции уровня сахара в крови он показал, что любые отклонения этого уровня от нормы включают процессы,.выравнивающие эти отклонения, что обеспечивает поддержание постоянства этой величины в организме. По этому же принципу регулируете: Постоянство температуры тела гомойотёрмцых животных и Другие параметры внутрен ней среды. Немецкий ученый Карл Людвиг и русский физиолог Ф. И. Цион обнаружили подоб ный (работающий по принципу отрицательной обратной связи) механизм, регулирую щий постоянство артериального давления в организме. Окончания чувствительной (депрессорного) нерва, локализованные в дуге аорты, при повышении давления кров: в этом сосуде посылают усиленные сигналы в ЦНС. Эти сигналы вызывают рефлекторно замедление сердцебиения и расширение артериол, что приводит к падению артериаль ного давления (т. е. к восстановлению его исходного уровня) . Затем в организме был< открыто большое количество подобных регуляторных механизмов. Значение в регуля ции движений обратных связей, т. е. сигналов, поступающих из работающих' мыши подчеркнул И. М. Сеченов. ' В ряде физиологических процессов был открыт механизм и положительной обратно1 связи/ благодаря которой процесс, возникнув, усиливается и поддерживает сам себя. Обратная связь — это связь на выходе системы. Она улавливает те или ины< отклонения, уже возникшие в состоянии системы, Основанные на этом регуляторньн механизмы работают по принципу «рассогласования». Деятельность их включаете) в тот момент, когда в состоянии системы уже наступают отклонения от заданной вели чины, т, е. когда 1 возникает рассогласование между заданной (необходимой) и факги чески возникшей величиной. Механизмы, работающие по этому принципу, широкс распространены в организме. Общий принцип работы подобных механизмов представле! П. К- Анохиным в схеме «функциональной системы» (см. рис. 243). Подобная схема однако, не является универсальной, т. к. в организме существуют регуляторные меха низмы, работающие на основе иного принципа. Сигналом к их деятельности служии отклонение от заданной величины не на выходе, а на входе системы, т. е. действие нг систему раздражителей, отличающихся от заданных параметров. В этом случае в основ) регуляторных реакций положен иной принцип, т. е. работа регулятора «по возмущению». На входе системы имеются приборы, улавливающие величину поступающего сигнала, нарушающего состояние системы. Если эта величина превышает допустимую н может вызвать нежелательные отклонения в состоянии системы, то в таком случае возникают команды, обеспечивающие нейтрализацию действия этих сигналов и сохранение стабильного состояния системы. Здесь происходит не восстановление уже нарушенного состояния системы, а предупреждение возможности таких нарушений. (Оба эти принципа сохранения стабильности системы отличаются друг от друга, как, скажем, средства тушения уже возникшего пожару отличаются от средств й мер предупреждения пожаров.) В любых физиологических регуляторных, защитных, компенсаторных реакциях имеет место взаимодействие обоих принципов и обоих механизмов регуляции, функционирующих как на выходе, так и на входе системы. Так, например, при воздействии на глаз струи .пыльного воздуха, которая может вызвать засорение глаза, срабатывают (как почти и везде) оба механизма. Мигательный рефлекс, закрывая глаз, предупреждает попадание пыли (это механизм,, работающий на входе системы «по возмущению»), а рефлекторное увеличение слезоотделения и промывание склеры ,и роговицы слезами удаляет уже попавшую пыль (механизм, работающий на выходе системы — «по рассогласованию»). В любой гомеостатической реакции можно наблюдать сочетание действия двух указанных механизмов, работающих на этих двух различных принципах. Для любой регуляторной реакции необходимо получение информации о состоянии системы, о величине поступающих сигналов, о возникающих при этом сдвигах в ее состоянии. Необходим также аппарат сличения параметров этих сдвигов или параметров поступающих сигналов с величиной нормальных для данной системы параметров. Кроме того, необходим аппарат, формирующий команды, предотвращающие эти Сдвиги. Действие этих команд осуществляется двумя путями: а) нормализацией уже возникших отклонений (механизмы, работающие «по рассогласованию»); б) предупреждением ежелательных эффектов входного (возмущающего) сигнала путем уменьшения силы игнала, предотвращения его действия или снижения чувствительности системы к дан- ому возмущающему воздействию (механизм, работающий «пО возмущению»).. Регу- 1яторные реакции осуществляются в организме нервной системой. |
|
|
 Скачать 7.39 Mb.
Скачать 7.39 Mb.