|
|
Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
Ацетилхолин. Ацетилхолин является уксуснокислым эфиром холина, т. е. относитс5 к простым эфирам. Он образуется при ацетилировании холина, причем этот процес» происходит, при участии фермента ацетилхолинтрансферазы. Особенностью, ацетилхоли на как медиатора является быстрое его разрушение после высвобождения из пресинапти ческих окончаний с помощью фермента -ацетилхолинэстеразы.
Ацетилхолин выполняет функцию медиатора, в синапсах,-образуемых возвратным} коллатералями аксонов двигательных нейронов спинного мозга на вставочных клетка; Реншоу, которые в свою очередь с помощью другого медиатора оказывают тормозяще» воздействие на мотонейроны.
Этот пример является хорошей иллюстрацией принципа Дейла, так как известно, что периферические окончания моторных аксонов активируют волокна скелетной мускулатуры с помощьк ацетилхЪлина. Возвратные коллатерали тех же аксонов в пределах ЦНС выделяют тот ж< медиатор.
Холинергическими являются и нейроны спинного мозга, ин нервируют не хром аффин- !ые клетки, а также преганглионарные нейроны, иннервирующие нервные клетки интра- 1уральныхи экстрамуральных ганглиев. Полагают, что холинергические нейроны имеются в составе ретикулярной формации среднего мозга, мозжечка, базальных ганглиях | коре.
Катехоламины. Три родственных в химическом отношении вещества: дофамин, юрадреналин и адреналин — являются производными тирозина и выполняют меди а торгую функцию не только в периферических, но и в центральных синапсах.
Дофаминергические нейроны находятся у млекопитающих главным образом в преде- Iах среднего мозга, образуя так называемую нигростриальную систему. Особенно ;ажную роль дофамин играет в полосатом теле, где обнаруживаются особенно большие юличества этого медиатора. Кроме того, дофаминергические нейроны имеются в гипо- ■аламусе. .
Норадренергические нейроны содержатся также в составе среднего мозга, моста гозга и продолговатого мозга. Аксоны норадренергических нейронов образуют восходящие пути, направляющиеся в гипоталамус, таламус, лимбические отделы коры и в мозжечок. Нисходящие волокна норадренергических нейронов иннервируют нервные клетки пинного мозга.
Катехоламины оказывают как возбуждающее, так и тормозящее действие а нейроны ЦНС. ,
Серотонин. Подобно катехоламинам, серотонин относится к группе, моноаминов, интезируется из аминокислоты триптофана. У млекопитающих серотонинергические ейроны локализуются главным образом в стволе мозга. Они входят в состав дорсального .медиального ядер шва продолговатого мозга;, моста и среднего мозга. Серотонинерги- еские нейроны распространяют влияния на новую кору, гиппокамп, бледный шар, [индалину, подбугровую область, стволовые структуры, кору мозжечка, спинной мозг.
Серотонин играет важную роль в нисходящем контроле активности спинного юзга и в гипоталамическом контроле температуры тела. Нарушения серотонинового бмена, возникающие при действии ряда фармакологических препаратов, могут вызы- ать галлюцинации. Нарушения функции серотонинергических синапсов наблюдаются :ри шизофрении и других психических расстройствах. Серотонин может вызывать возбуждающее и тормозящее действие в зависимости от свойств рецепторов постсинап- ической мембраны.
Нейтральные аминокислоты. Две основные дикарбоксильные кислотыL-глутамат и .-аспартат находятся в большом количестве в ЦНС и могут выполнять функцию медиа- оров. •
. L-глутаминовая кислота представляет собой дикарбоновую аминокислоту, входящую в состав многих белков и пептидов. Она плохо проходит через гематоэнцефали- еский барьер и поэтому не поступает в мозг из крови, образуясь в самой нервной кани (главным образом из глюкозы) . В ЦНС млекопитающих глутамат обнаруживается высоких концентрациях. По-видимому, он является одним из самых распространенных [едиаторов в центральных синапсах позвоночных животных. Полагают, что его функция вязана главным образом с синаптической передачей возбуждения.
Глутамат исчезает из синаптической щели вследствие захвата его нервными и гли- льными клетками и пресинаптическими окончаниями. Глутамат принимает, участие1 ряде важных метаболических процессов и входит в цикл синтеза 7-аминомасляной ислоты. Сходное с ним действие оказывает на центральные нейроны аспартат. ,
Кислые аминокислоты. К этой группе аминокислот относятся ГАМК и глицин.
ГАМК представляет собой продукт декарбоксилирования L-глутаминовой кислоты. Эта реак- ия катализируется декарбоксилазой глутаминовой кислоты. Отмечено значительное совпадение окализации этого фермента и ГАМК в пределах ЦНС. Другой фермент нервной ткани — трансами- аза — катализирует перенос аминогруппы ГАМК на а-кетоглутаровую кислоту, в результате чего оследняя превращается в семиальдегид янтарной кислоты.
• ГАМК содержится в нейронах спинного и головного мозга. При ее апНликнци к различным нейронам ЦНС почти всегда возникает тормозной эффект, вследствие чёг ГАМК рассматривают как наиболее распространенный медиатор1 синаптическото торм*с жения. Так, тормозное действие ГАМК было продемонстрировано на клетках кор больших полушарий, нейронах ствола мозга, двигательных нейронах спинного мозг; ГАМК выполняет функцию медиатора при осуществлении как постсинаптйческого, та и Пресинаптического торможения (см. ниже). ,
Медиаторная функция глицина ограничивается главным образом спинным Мозго* где это вещество выполняет роль меДиатора постсинаитического торможения.
Так же как нейтральные аминокислоты, ГАМК и глицин после своего освобождени пресинаптическими окончаниями удаляются из синаптической щели путем захват нервными й глиальными клетками. ' ' - .
Полипептиды. В последние годы показано, что в синапсах ЦНС медиаторную фуш цию могут выполнять некоторые полипептиды. К таким полипептидам относятся et щество Р, гипоталамические нейрогормоны, энкефалин и др. 1
Под названием «вещество Р» подразумевается группа агентов, впервые экстрагйр< ванных из кишечника. Эти полипептиды обнаруживаются во многих частях ЦНС. Oco6ei но высока их концентрация в области черного вещества. Наличие вещества Р в задни корешках спинного мозга позволяет предполагать, что око может служить мед и а тор о в синапсах, образуемых центральными окончаниями аксонов некоторых пёрвичнь афферентных нейронов. Действительно, вещество Р оказывает возбуждающее действр на определенные нейроны спинного мозга.
Медиаторная роль других нейропептидов выяснена еще меньше; Специфические рецепторы мембраны. Для химической передачи в синапсах необхс дймо существование особых мембранных рецепторов, с которыми реагируют химически медиаторы." Результатом этого взаимодействия является специфическое 'изменен!' свойств постсинаптической мембраны, приводящее к возбуждению или торможен и постсинаптической клетки. '
Роль мембранных рецепторов играют белковые молекулы, обладающие способ ностью «узнавать» специфические для них вещества й вступать с ними в реакцию. Bej ковые молекулы подвергаются конформационным изменениям, вследствие чего происхе дит активация специальных ионных каналов мембраны (ионофоров). В результате этог процесса изменяется ионная проницаемость мембраны, что в свою очередь изменяв мембранную проводимость и приводит к уменьшению или увеличению трансмембран не разности потенциалов—деполяризации или гиперполяризации. • '
В настоящее время стало очевидным, что рецепторы мембраны довольно быстр обновляются/Они синтезируются, Вероятно, в энДоплазматичееком р ет и кул ум е, в кл юче i ном в аппарат Гольджи, и оттуда переносятся к поверхности нервной клетки ii включают с я- в ее мембрану. Весь процесс занимает несколько часов.
Один и тот жё медиатор может вступать- в реакцию с различными рецепторам постсинаптической мембраны и: вызывать противоположные эффекты. Так, в нейрона ЦНС обнаружены мускариновые и никотиновые холинорецепторы, воздействуя на кот( рые ацетилхолин вызывает различные изменения проницаемости постсинаптическо мембраны. Показано существование различных рецёпторов к; катехоламинам. Накаг ливается .все больше данных в пользу существования различных рецепторе к аминокислотам. ; • . ■ . ■
Способность одного и. того же медиатора вызывать разнонаправленные из мене ни пройицаемости постсинаптической мембраны является причиной того, что одни и fe ж медиаторы могут или возбуждать, или тормозить различные нервные клетки. В те случаях, когда влияние химического медиатора более однотипно, как, например, в случа ГАМК и глицина, действие которых почти всегда приводит к увеличению хлорной пронр цаемости мембраны, функциональный эффект оказывается однозначным (тормозны в случае указанных аминокислот).
Возбуждающий синапс
Энстраклеточкая ? среДа
i fc
R;
—Г "'1—
Внутриклеточная среда
4ic. 66- Эквивалентная электрическая схема озбуждающего синаптического входа.
несинаптическая область
'м—емкость мембраны; J?m> — сопротивление ембраны; Е^-—электродвижущая сила мембра- ы; Е^ — электродвижущая сила синаптического онтакта; Rs — сопротивление синаптического он такта. ,
мВ м
: 3 4 не
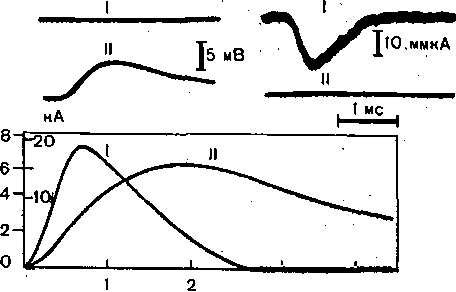
Рис. 67. Возбуждающий постсинаптический потенциал (ВПСП) и соответствующийему постсинаптический ток в мотонейроне спинного мозга кошки.
а: I—ток при отсутствии фиксации потенциала; II — ВПСП; б: I — ток, II —потенциал во время фиксации; в — график динамики постсинаптического тока (I) и ВПСП (II).
ВОЗБУЖДЕНИЕ В центральной НЕРВНОЙ системеХотя один й тот же химический медиатор, действуя на разные рецепторы постси- аптичеекой мембраны в различных нервных, клетках может вызывать как возбуждающие, так и тормозные процессы, в ЦНС позвоночных можно выделить синапсы, которые ыполняют однозначную функцию — возбуждения (возбуждающие синапсы) (рис. 66). 'ак, центральные отростки первичных афферентных нейронов всегда оказывают возбуж- ающее действие на нейроны спинного мозга. Другим примером возбуждающего ействия у позвоночных является мотонейрон, активирующий не только мышцы, но вставочные клетки Реншоу спинного мозга. В возбуждающих синапсах медиатор, высвобождаемый пресинаптическим оконча- ием, вызывает развитие локального процесса деполяризации, обозначаемого как озбуждающий постсинаптический потенциал (ВПСП). Указанное название подчерки- ает тот факт, что ВПСП развивается в постсинаптической мембране. В ЦНС млекопитающих ВПСП наиболее подробно изучены в спинальных мотонейро- :ах, где имеется возможность избирательной активации однородных по составу фферентных волокон, которые образуют синапсы непосредственно на мотонейронах. Иго позволяет изучать, моносинаптические эффекты, не связанные с вовлечением в прочее возбуждения вставочных нейронов (рис. 67). Амплитуда ВПСП зависит от исходного уровня мембранного потенциала. Смещение мембран- ого потенциала до величин, близких к нулю, обычно приводит к извращению (реверсии) знака >ПСП, т. е. суммарный постсинаптический ток в, этих условиях течет в обратном направлении, 'то означает, что активированная возбуждающим медиатором постсинаптическая мембрана ста- овится проницаемой не только для ионов натрия, но и для некоторых других ионов, содержащихся нутри и снаружи клетки. Опыты с введением внутрь нейрона С1 показали, что при этом амплитуда 1ПСП не изменяется. По-видимому, возникновение ВПСП связано с одновременным увеличением роницаемости постсинаптической мембраны для Ма 2+ и К +, а также, возможно, Са 2+. Увеличе- ие калиевой проницаемости приводит к уменьшению деполяризации, которая могла бы возникнуть а счет увеличения только натриевой или натриевой и кальциевой проницаемости. Деполяризация нервной клетки в результате действия возбуждающего медиатора ВПСП) может быть достигнута не только за счет увеличения проницаемости ее мембра- :ы для Na + (или Са 2+), но и за счет уменьшения проницаемости для К +-
•Важным показателем эффективности синаптического возбуждения нервной клеть является способность возбуждающих синапсов вызывать возникновение потенции действия. Необходимым условием для генерации потенциала действия является сниж< ние трансмембранной разности потенциалов постсинаптической мембраны до определе] ного критического уровня.
Условия возникновения потенциала действия в нервной клетке под влиянием ci наптического возбуждения в значительной степени обусловлены неодинаковой электр] ческой возбудимостью различных участков мембраны и пространственным распределен] ем различных возбуждающих синапсов. В большинстве центральных нейрож потенциал действия возникает в специальной низкопороговой области (обычно Э1 зона аксонного холмика), откуда он распространяется по аксону и на мембрану сосе, них участков клетки. Указанный способ синаптического возбуждения нейрона оче! важен для его интегративной функции, т. е. способности суммировать влияния, поступ ющие на нейрон по разным синаптическим путям.
ТОРМОДЕНИЕ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
Одним из фундаментальных свойств ЦНС является способность к торможенш Явление центрального торможения было открыто в 1863 г. И. М, Сеченовым, обнаружи шим возникновение торможения спинальных центров лягушки при раздражении структ; среднего мозга. В дальнейшем была выявлена возможность торможения спинальнь реакций не только при раздражении-надсегментарных образований, но и нервов прот воположной стороны тела. Это открытие позволило подойти к установлению реципро ных отношений между процессами возбуждения и торможения в ЦНС. Реципрокнь характер возбуждающих и тормозных влияний в спинном мозге показан ученике И. М. Сеченова Н. Е. Введенским и подробно проанализирован английским нейрофизи логом Ч, Шеррингтоном.
Важным шагом в выяснении природы центрального торможения оказалось выявл ние самостоятельного значения торможения длд работы нервных центров. Торможен] нельзя свести ни к утомлению нервных центров, ни к их перевозбуждению или катод ческой депрессии нервных клеток. Торможение — самостоятельный нервный процес вызываемый возбуждением и проявляющийся в подавлении другого возбуждения. В о личие от процесса возбуждения, который может проявляться в двух основных формах распространяющихся потенциалов действия и локальных потенциалов, торможен; может развиваться только в форме локального процесса и поэтому всегда связа: с существованием специфических тормозных синапсов (рис. 68).
Функция тормозных синапсов однозначна (они всегда вызывают только тормож ние), образующие их пресинаптические окончания Относятся к аксонам так называемь тормозных нейронов, угнетающих активность всех нервных клеток, которые они иннерв руют. Примером тормозных нейронов в спинном мозге являются вставочные нейроь Реншоу, в головном мозге — грушевидные нейроны (нейроны Пуркинье) коры мозжечк
С помощью микроэлектродов установлено, что синаптич'еское торможение мож вести к подавлению активности нейрона, имеющего тормозной синапс, вследствие изм нения свойств постсинаптической мембраны нейрона (так называемое постсинаптическ торможение) или в результате уменьшения эффективности действия возбуждающ: синапсов еще на пресинапТическом уровне (так называемое пресинаптическое торм жен'ие). Особенно широкое распространение в ' ЦНС имеет постсинаптическ торможение.
Постсинаптическое торможение. Медиатор, выделяемый пресинаптическими оконч ниями тормозных синапсов, изменяет свойства постсинаптической мембраны таким обр зом, что способность нервной клетки генерировать процессы возбуждения (ВПСП и, потенциал действия) подавляется. Поэтому данное явление принято обозначать к постсинаптическое торможение, а лежащее в его основе изменение в постсинаптическ мембране — тормозной постсинаптический потенциал,(777СП).
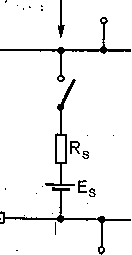
Тормозящий :' синапс 5 ''' -'' ' ' : Э я стр аил ето ч н ая среда есинаптическая область  0,1 нВ 1 мс Внутриклеточная среда
1С. 68. Эквивалентная электрическая схема рмозного синаптического входа.
л— емкость мембраны; Rm, Rj—сопротивление мбраны; Rs— сопротивление синаптического нтакта; Ем -— электродвижущая сила мембраны, ;—электродвижущая сила синаптического кон- кта.
Рис. 69. Тормозной постсинаптический потенциал (ТПСП) в мотонейронах спинного мозга кошки.
а — ТПСП, вызванный раздражением нерва; б — ТПСП, вызванный раздражением вставочного нейрона.
Специфика тормозных синаптических эффектов была впервые наиболее подробно ;учена на мотонейронах млекопитающих, а в дальнейшем — на многих нейронах мозга, ;лючая нервные клетки мозгового ствола, гиппокампа и коры. В мотонёйронах спинного мозга возникновение ТПСП в ответ на раздражение эффектных волокон, идущих от мышц-антагонистов, обязательно связано с включением тормозной процесс дополнительного звена —специального вставочного тормозного ^йрона, аксональные окончания которого выделяют медиатор (вероятнее всего это 1ицин), вызывающий развитие ТПСП в постсинаптической мембране. Пример ТПСП мотонейроне спинного мозга кошки показан на рис. 69. Как видно на этом рисунке, >ёменноё течение ТПСП почти совпадает с временным течением ВПСП. И для ВПСП, для ТПСП характерна более быстрая фаза нарастания и более длительная, убывающая > экспоненте фаза спада. ТПСП, возникающие при раздражении мышечных нервов, }жно рассматривать как результат почти синхронного вовлечения совокупности тормозах нейронов. .ТПСП, вызываемые прямым микроэлектродным раздражением одного >рмозного нейрона, имеют сходные временные характеристики, но значительно мень- ую величину. Первоначально было сделано заключение, что торможение всегда развивается результате гиперполяризации постсинаптической мембраны, так как тормозной медиа- >р увеличивает ее проницаемость для К +- В дальнейшем было установлено, что постси- штическоё торможение не обязательно сопровождается гиперполяризацией мембраны, iK как более важное значение имеют лежащие в основе ТПСП сложные изменения >нной проводимости постсинаптической мембраны. ТПСП обнаруживает очень высокую чувствительность к сдвигам мембранного потенциала, !еличиваясь при деполяризации и уменьшаясь при гиперполяризации. Когда последняя приводит увеличению' мембранного потенциала до 80 мВ, ТПСП превращается в деполяризационный вет. Однако и в этом случае его тормозящее действие сохраняется. . Извращение ТПСП объясняется тем, что тормозной медиатор повышает проницаемость >стсинаптической мембраны для!СР. В нормальных условиях концентрации CP во внеклеточной 1еде превышает его содержание в нейроплазме. Во время развития ТПСП отрицательно заряжен- >ig ионы хлора устремляются внутрь клетки, увеличивая трансмембранную разность потенциалов, огда концентрация CP-в нейроплазме превышает его содержание в наружной среде, тормозной гдиатор приводит к движению CP из клетки наружу, что приводит к ее деполяризации в резуль- iTe потери отрицательных зарядов. Таким образом, тормозная постсинаптическая мембрана мото- ?йронов и других нейронов ЦНС действует как образование селективное к CP, что, вероятно, 5условлено наличием положительных зарядов в стенках ионных каналов мембраны.
Физический смысл ТПСП всегда остается неизменным, он стремится сдвинуть мембраннь потенциал в сторону, противоположную той, которая необходима для развития возбуждающе! эффекта.
Учитывая природу тормозного процесса, можно сделать вывод, что эффективность тормознь синапсов во многом зависит от их локализации на поверхности клетки. Тормозной эффект тем бол' значителен, чем ближе тормозной синапс расположен к месту генерации потенциала действи Вследствие этого тормозные синапсы локализованы главным образом на теле нервных клет< вблизи от триггерной зоны аксонного холмика.
Поскольку функция тормозных синапсов заключается именно в подавлении или ограничен процессов возбуждения, развивающихся в постсинаптической мембране, важно рассмотре' особенности взаимодействия. возбуждающих и тормозных постсинаптических эффекта
В клетках ЦНС, получающих как возбуждающие, так и тормозные синаптические вход| их взаимодействие может быть рассмотрено на примере суммации ВПСП и ТПСП. Исследовани проведенные с помощью усреднения постсинаптических потенциалов на вычислительной машин показали, что суммация ВПСП и ТПСП обычно имеет нелинейный характер (рис. 70).
Поэтому суммарная реакция нервной клетки на сочетанную активацию взаимодействующи входов значительно меньше алгебраической суммы обоих потенциалов. Наибольшая степе! нелинейности наблюдается при совмещении начальных фаз ВПСП и ТПСП, т. е. в момент, ког; лежащие в их основе изменения проводимости достигают максимума. Это полностью согласует! с тем, что эффект постсинаптического торможения обусловлен в первую очередь повышением пров димости постсинаптической мембраны.
Пресинаптическое торможение. Синаптическое торможение, приводящее к умен! шению эффективности возбуждающих синаптических влияний, может развиваться и только на уровне постсинаптической мембраны (как это было рассмотрено выше), н еще в пресинаптическом звене путем: угнетения процесса высвобождения медиатор возбуждающими нервными окончаниями. В этом случае свойства постсинаптическо мембраны не подвергаются каким бы то ни было изменениям.. ;
Пресинаптическое торможение обнаружено в различных отделах ЦНС: Наиболе часто оно выявляется в структурах мозгового'ствола и особенно в спинном мозп
Так же как и постсинаптическое, пресинаптическое торможение осуществляете посредством специальных тормозных вставочных нейронов.
Рис. 70. Суммация возбудительного и тормо; ного постсинаптических потенциалов.

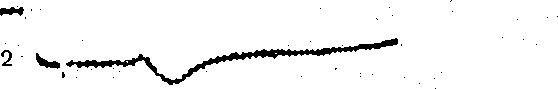
1 —ВПСП; 2- ТПСП'; 3 — суммация ВПСП ТПСП'; 4 — суммация ВПСП и ТПСП"; 5- ТПСП".
Рис. 71. СхемаОрганизации синапсов, участв} ющих в пресинаптическом торможении.
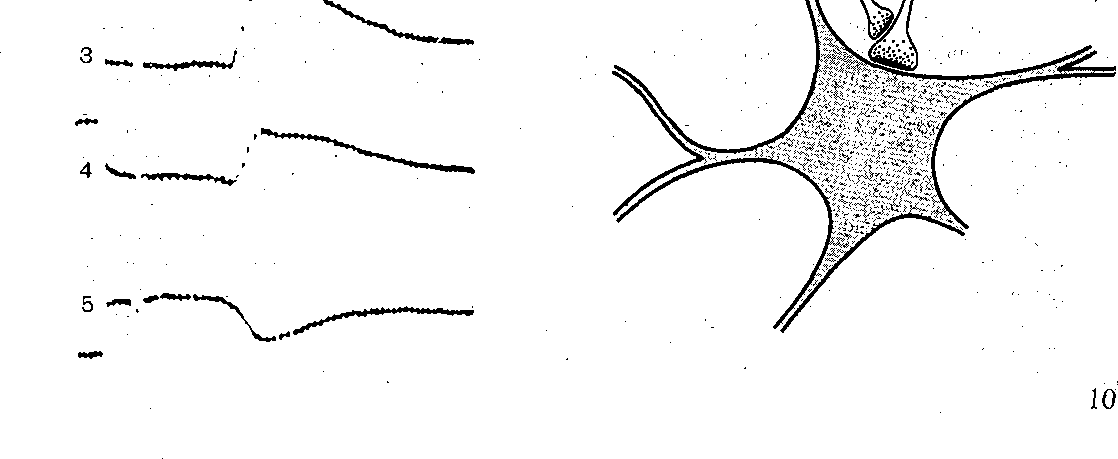
Структурной основой пресинаптического торможения являются аксоаксонные инапсы, образованные окончаниями аксонов тормозных вставочных нейронов и аксо- альными окончаниями возбуждающих нейронов. В этом случае окончание аксона ормозного I нейрона является пресинаптическим по отношению к возбуждающему кончанию, которое в свою очередь будучи постсинаптическим по отношению ктормозно- :у окончанию, является пресинаптическим по отношению к активируемой им нервной летке (рис. 71). Импульсы в пресинаптическом тормозном аксоне высвобождают медиа- ор (в спинном мозге это вероятнее всего у-аминомасляная кислота)у который вызывает еполяризацию возбуждающих окончаний за счет увеличения проницаемости их мембра- ы для С1 Предполагается, что указанная деполяризация вызывает уменьшение мплитуды потенциала действия, приходящего в возбуждающее окончание, что в свою чередь уменьшает количество высвобождаемого им медиатора, вследствие чего ампли- уда возбуждающего постсинаптического потенциала падает.
Другим механизмом пресинаптического торможения может быть уменьшение входящего нутрь потока Са2+, воздействующего на электросекреторную связь. И в этом случае пресинаптиче- кое торможение приводит к уменьшению числа квантов медиатора, высвобождаемого возбуждающим пресинаптическим окончанием.
У млекопитающих продолжительность пресинаптического торможения значительно ревосходит продолжительность постсинаптического торможения. Большая длитель- ость пресинаптического торможения^ по-видимому, обусловлена ритмической актив- остью тормозных вставочных нейронов. ;..■'. : ■
Пресинаптическое торможение особенно эффективно при обработке информации, уступающей к нейрону по различным, пресинаптическим путям. В этом случае возбуж- ,ение, поступающее по одному из синаптических входов, может быть избирательно меньшено или даже полностью подавлено при отсутствии влияния на другие входы. 1одобного результата нельзя достичь путем воздействия на проводимость постсинапти- 1еской мембраны, как это имеет место при постсинаптнческом торможении, влияющем ia всю нервную клетку.
ИНТЕГРАЦИЯ СИНАПТИЧЕСКИХ ВЛИЯНИЙ
Каждая клетка ЦНС имеет множество синаптических контактов с различными ней- он.ами. Так, на одной клетке Пуркинье.коры мозжечка насчитывают до 200.000 синапсов, исло синапсов на мотонейронах млекопитающих составляет от 10 000 до 20.000.
Дивергенция. Способность нейрона устанавливать многочисленные ..синаптические вязи с различными нервными,-клетками носит название дивергенции. Например, цент- ильные окончания аксонов первичного афферентного нейрона образуют, синапсы на шогих мотонейронах-синергистах, на вставочных нейронах, осуществляющих торможе- ше мотонейронов-антагонистов, и на клетках,, дающих .начало дорсальному спиноце- «ебеллярному восходящему тракту. Благодаря процессу дивергенции одна и та же 1ервная клетка может участвовать в различных нервных реакциях и контролировать юльшое число других нейронов, а каждый, нейрон может обеспечивать широкое 1ерераспределение импульсов, что приводит к иррадиации возбуждения. •■•'
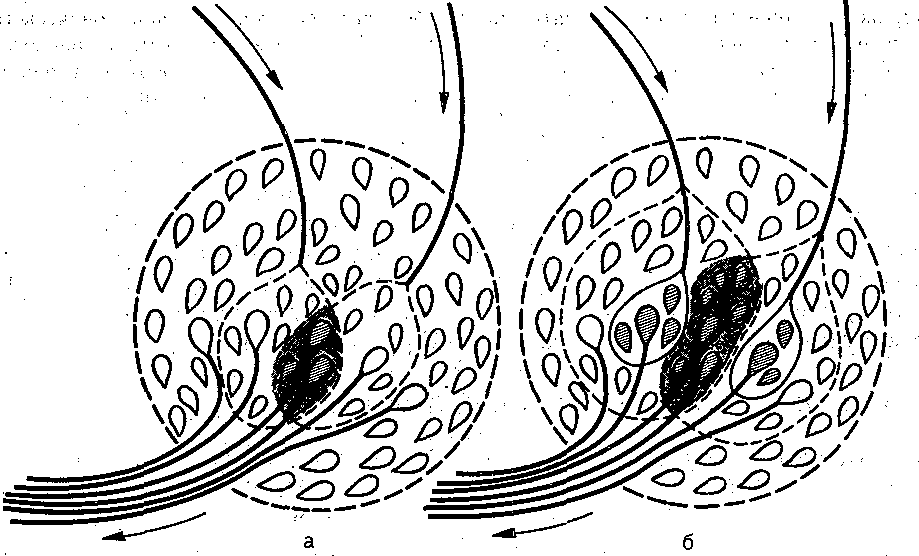
Конвергенция. Схождение различных путей проведения нервных импульсов к одной { той же нервной клетке носит название конвергенции. Простейшим примером конвер- ■енции служит факт Получения, каждым мотонейроном импульсов от совокупности тервичных аф.ферентных;нейронов. На рис. 72 схематически показана конвергенция цент- >альных окончаний первичных афферентных волокон на одном и том же мотонейроне \ продемонстрирован конкретный пример развития ВПСП в мотонейроне 3 различными 1ресинаптическими элементами. . , '
Если в приведенном выше примере конвергенция ограничивается эффектами, вызываемыми однотипными нейронами (все они относятся к сенсорным нейронам, проводя-
Объяснение в
Схема конвергенции.
Рис. 72. тексте.
Рис. 73. Линейная суммация ВПСП Объяснение в тексте.
щим в спинной мозг импульсы от мышечных рецепторов растяжения), то значительна часть нервных клеток ЦНС имеет синапсы с нейронами различного типа, обеспечивак щими конвергенцию влияний из разных источников. Например, к мотонейронам спинног мозга, кроме первичных афферентных волокон, конвергируют волокна различных нисхс дящих трактов, берущих начало в супраспинальных' и собственно спинальных центра: аксоны возбуждающих и тормозных вставочных нейронов. Поэтому мотонейроны ра( сматриваются как общий конечный путь многочисленных нервных структур, связанны с регуляцией моторной функции ЦНС. Принцип общего конечного пути был введен в фр зиологию нервной системы Ч. Шеррингтоном. Он показывает, каким образом одна и та ж конечная реакция, проявляющаяся активацией определенной группы мотонейроно! может быть получена при раздражении различных нервных структур. Данный принци имеет первостепенное значение для анализа рефлекторной деятельности (см. ниже нервной системы. Синаптическое взаимодействие. Конвергенция различных синаптических входов н одной нервной клетке обеспечивает возможность их взаимодействия. Так. при активаци различных возбуждающих синапсов происходит пространственная суммация ВПСГ. Пространственная суммация возбуждающих синаптических влияний имеет важно значение для возникновения импульсной активности в нервной клетке, так как денолнр! зации, создаваемой одним синаптическим входом,.часто бывает недостаточно для достр жения порогового уровня и генерации потенциала действия. Пространственная и вре менная суммация ВПСП способна приводить к длительной деполяризация постсинаптк ческой мембраны, что обеспечивает возникновение ритмической импульсной активност нервной клетки. / Возникающие при активации различных синапсов ВПСП могут. суммироваться линейнс При линейной 'суммации общая деполяризация равна арифметической . сумме деполяризаций создаваемых каждым входом в отдельности.. Возможна и нелинейная суммация, когда . обща деполяризация.нейрона меньше, арифметической суммы деполяризаций, создаваемых каждым воз буждающим синаптическим входом. Пример линейной суммации ВПСП, возникающих в одно и той же нервной клетке при раздражении разных пресинаптических путей, показан на рис] 73. Линейный характер суммации наблюдается в том случае, когда взаимодействующие возбуж дающие синапсы, конвергирующие на данном нейроне, расположены на таком расстоянии дру от друга, когда повышение проводимости постсинаптической мембраны, развивающееся под влия нием возбуждающего медиатора, не оказывает шунтирующего влияния на соседний вход. Наоборот при достаточно близкой локализации взаимодействующих синаптических входов нервной клетк| увеличение проводимости постсинаптической мембраны будет шунтировать и, следовательно, умень to:
[ать деполяризацию, создаваемуфхоседним входом. Как отмечалось выше^суммация возбуждаю- 1их и тормозных постсинаптических потенциалов обычно развивается нелинейно. Чем выше степень глинейности; тем сильнее выражен'тормозной эффект!
Кроме взаимодействия непосредственно на постсинаптической мембране, различные инаптическиё влияния могут взаимодействовать еще и на пути к нервной клетке. Одним з примеров, такого взаимодействия может служить рассмотренное выше пресинапти- еское торможение. Кроме того, на пресинаптическом уровне может развиваться и провес облегчения. Такое пресинаптическое или гетеросинаптическое облегчение заключа- гся в увеличении эффективности одного синаптического возбуждающего входа результате .активации другого; когда облегчение развивается не на уровне постсинап- ической мемДраны нейрона, а вследствие взаимодействия на пути к нему.
Сами нервные клетки нередко обладают возможностью регулировать величину оступающих к ним сигналов. Такой механизм, получивший название обратной связи, аключается в том, что коллатерали аксонов нервной клетки могут устанавливать си- аптические контакты со специальными вставочными нейронами, роль которых заклю- ается в воздействии на нейроны или аксональные.окончания путей, конвергирующих а нервной клетке, посылающей эти аксонные коллатерали. Так, например, возникнове- ие импульса в мотонейроне млекопитающих не только активирует мышечные волокна, о и через коллатерали возбуждает специальные тормозные клетки Реншоу. Аксоны леток Реншоу в свою очередь устанавливают синаптические связи с мотонейронами. 1оэтому, чем сильнее импульсация мотонейрона, тем больше активируются Клетки 'еншоу и тем значительнее они тормозят мотонейроны, уменьшая частоту их импульса- ии (так называемое возвратное торможение).
Благодаря наличию обратных связей, степень возбуждения нейронов различных ервных центров может строго согласовываться как с интенсивностью приходящих к ним озбуждающих влияний, гак и с интенсивностью импульсации на выходе нейронов и, сле- оватедьно, с интенсивностью развиваемого рабочего эффекта. Так, мотонейроны полу- ают информацию о сокращениях мышцы от сухожильных и мышечных рецепторов, ти импульсы, сигнализирующие о состоянии двигательного аппарата, позволяют орригировать активность этих мотонейронов. Афферентные импульсы, поступающие от эсудов, органов дыхания, пищеварения й выделения, постоянно корригируют деятель- ость нейронов, участвующих в поддержании уровня артериального давления и регу- яции Других вегетативных функций.
РЕФЛЕКТОРНАЯ ДЕЯТЕЛЬНОСТЬ ЦНС
Взаимодействие нервных клеток составляет основу целенаправленной деятельности ервной системы и прежде всего осуществления рефлекторных актов.
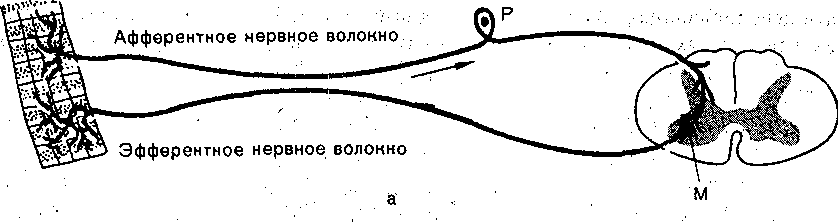
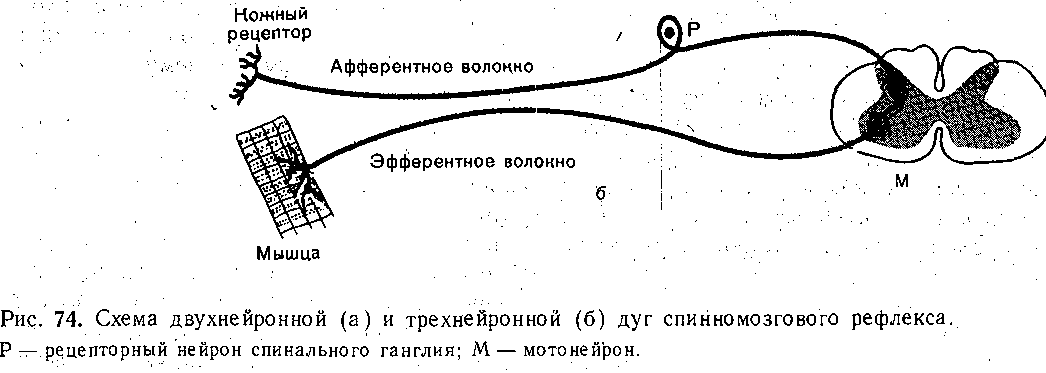
Как уже было сказано, принцип рефлекторной (отражательной) деятельности ервной системы был выдвинут еще в XVII в. французским философом и математиком '. Декартом. Сам термин «рефлекс» был предложен в XVIII в. чешским физиологом 1рохазкой. Весь последующий ход изучения деятельности нервной системы убедительно оказал, что ее ответы на различные раздражения протекают по рефлекторному принци- у. Рефлекторную природу психической деятельности обосновал И. М. Сеченов. . V Рефлекторная дуга. Основой рефлекторного ответа является так называемая дуга ефлекса •• •• комплекс специфически организованных нервных элементов, взаимодействие оторых необходимо, для осуществления рефлекторного акта. Рефлекторная дуга состоит ;з афферентной, центральной и эфферентных частей, связанных между собой с помощью инаптических соединений (рис. 74).
Афферентная часть представлена теми нервными элементами, которые формируют : проводят в центральном направлении нервные импульсы, необходимые для деятельности •сей рефлекторной дуги. Поскольку возникновение афферентных импульсов связано активацией специфических рецепторов, совокупность рецепторов, раздражение которых
вызывает определенный рефлекс, называют рецептивным полем рефлекса. Следуе отметить^ что раздражение одних и тех же рецепторов не всегда вызывает один и то же тип рефлекторного ответа, а могут отмечаться различные рефлексы в зависимост от того, к каким центральным структурам импульсы поступают по первичным афферент ным нейронам. Кроме того, в рецептивном поле одного рефлекса могут находитьс и различные по функции рецепторы. Так, сгибательный рефлекс может вызыватьс: раздражением тактильных'рецепторов кожи или мышечных рецепторов.
Поступающие по афферентным путям нервные импульсы с помощью синаптически. переключений активируют различные нейроны. ЦНС. Часть афферентных импульсоЕ необходимых для возникновения рефлекторного ответа, переключается также на нейрош восходящих трактов и отражается в сознании. Однако многие рефлексы возникают и бе участия сферы сознания, так как для их осуществления достаточно участие подкорковы зон ЦНС. Именно поэтому многие рефлекторные акты могут сохраняться даже поел разрушения большей части ЦНС. Рефлекторная деятельность может осуществлятьс и одним изолированным сегментом спинного мозга, выделенным из организма и перфу зируёмым искусственным путем.
Моно- йполисинаптические рефлексы. Центральная часть рефлекса может включат несколько последовательных нейронов,-соединенных синаптическими контактами. Тогд рефлекс носит название полисинаптического. В простейшем случае импульсы, поступа ющие в ЦНС по афферентному пути, переключаются непосредственно на эфферентны нейрон. Поскольку в пределах ЦНС рефлекторная 'дуга такого рефлекса имее только один синапс (например, синапс между центральными окончаниями мышечны афферентов и мотонейронами), он носит1 название моносинаптйческого. Примером монс синаптического рефлекса является сухожильный рефлекс,- или рефлекс растяжение
В результате возникновения импульсов в эфферентных нейронах происходи активация эфферентной части рефлекса и его реализация. Рефлексы очень многообразш и их исполнительная часть включает различные органы и системы организма. В большин стве своем рефлексы служат для защиты организма и приспособления его к изменения! окружающей и внутренней среды. С их помощью адекватно координируются нелроиз вольные акты организма. Секреция желез, движения внутренних органов, реакци
'ис. 75. Окклюзия. Объяснение в тексте.
ердца и сосудов, скелетной мускулатуры тонко контролируются координированными ефлекторными актами.
Большинство детальных сведений о рефлекторных актах было получено благодаря зучению мышечных ответов, поскольку последние дают объективную и хорошо иамеря- мую оценку рефлекса. Примером двигательного рефлекса у человека является колен- ый рефлекс. Поскольку рефлекс не может быть вызван после того, как соответствующий .ентр разрушен или поврежден, наличие или отсутствие определенных рефлексов имеет ажное диагностическое значение. Так, наличие коленного рефлекса свидетельствует сохранности моторных центров поясничного сегмента спинного мозга. Рефлекторное окращение зрачка в ответ на освещение показывает, что ядро III черепного нерва соответствующие зрительные и эфферентные пути являются интактными.
Характер рефлекса в значительной степени зависит от интенсивности раздражения числа активируемых рецепторов. Усиление раздражения приводит к расширению ецептивного поля рефлекса, в результате чего вовлекается большее число центральных ейронов. Указанное явление называется иррадиацией возбуждения. Процесс иррадиа- ии в значительной степени зависит от того, что отдельные афферентные нейроны ызывают подпороговую деполяризацию центральных;нейронов. При увеличении числа ктивированных афферентных нейронов в результате п|роцессов пространственной уммации в большем числе центральных нейронов синаптическое возбуждение достигает орога и приводит к их импульсной активности.
Усиление раздражения вызывает также возрастание частоты импульсации в аффе- ентных волокнах, что в свою очередь увеличивает ответы центральных нейронов следствие временной суммации возбуждающих синаптических влияний.
Взаимодействие рефлексов. Различные рефлекторные реакции могут взаимодейство- ать между собой. Примером такого взаимодействия является феномен доминанты .. А.' Ухтомского. Образование в ЦНС центра повышенной возбудимости приводит тому, что раздражение самых, различных рецептивных полей начинает вызывать ефлекторный ответ, характерный для деятельности этой доминантной области.
Доминантный очаг в ЦНС может возникать под влиянием разных, факторов, например результате гормональных воздействий. В частности, в период спаривания половые гормоны звышают возбудимость моторных центров шейного утолщения спинного мозга амфибий и любое раздражение кожи начинает вызывать вместо обычного рефлекса усиление тоническог обнимательного рефлекса. После кастрации эти рефлексы угасают, но вновь восстанавливаютс при введении полового гормона тестостерона. Показательно, что усиление активности нейроне моторных центров развивается даже при изоляции спинного мозга и добавлении тестостерон в .перфузирующий раствор.
Взаимодействие рефлексов может проявляться как во взаимном облегчении (сум мация), так и угнетении (окклюзия). Последнее явление выражается в том, чт суммарный результат оказывается значительно меньшим, чем сумма взаимодействующи реакций. Согласно Ч. Шеррингтону (рис. 75), явление окклюзии объясняется перекры тием синаптических полей, образуемых афферентными частями взаимодействующих реф лексов. Поэтому при одновременном поступлении двух афферентных влияний ВПС1 вызывается каждым из них отчасти в одних и тех же мотонейронах.
ОБЪЕДИНЕНИЕ НЕЙРОНОВ В НЕРВНЫЙ ЦЕНТР
В результате интегративной деятельности нейронов ЦНС, в основе которой лежа процессы суммации возбуждающих и тормозных постсинантических потенциалов, воз никающих в разных участках поверхностной мембраны, в нервной клетке происходи генерация импульсов или команд, регулирующих, деятельность других нейронов ил органов. В этом смысле работа индивидуального нейрона в какой-то степени подобн работе нервной системы в целом. Однако сложность и многообразие задач, выполняемы нервной системой, приводит к высокой специализации отдельных нейронов, и не тольк отдельных нейронов, но и нейронных объединений — нервных Центров, деятельност которых связана с осуществлением различных функций. Функционирование ЦНС в цело! осуществляется благодаря деятельности значительного числа таких центров, представ ляющих собой ансамбли нервных клеток, объединенных с помощью синаптически контактов и отличающихся огромным разнообразием и сложностью внутренних и внен; них связей.
В соответствии с выполняемой функцией можно выделить различные чувствительны центры, центры вегетативных функций, двигательные, центры, центры психически, функций и т. д. Различные нервные центры характеризуются определенной топографие своего расположения в пределах ЦНС и более подробно будут рассмотрен) в следующей главе.
|
|
|
 Скачать 7.39 Mb.
Скачать 7.39 Mb.