Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
 Скачать 7.39 Mb. Скачать 7.39 Mb.
|

|
Глава 4 ПРОВЕДЕНИЕ НЕРВНОГО ИМПУЛЬСА И НЕРВНО-МЫШЕЧНАЯ ПЕРЕДАЧА ПРОВЕДЕНИЕ НЕРВНОГО ИМПУЛЬСА СТРУКТУРА НЕРВНЫХ ВОЛОКОН ' Проведение нервных импульсов является специализированной функцией нервньи волокон, т. е. отростков нервных клеток. . . Нервные волокна разделяют на мякотные, или миелйнизированные, и безмякотные немиелинизированные. Мякотные, чувствительные и двигательные волокна входят е состав нервов, снабжающих органы чувств и скелетную м'ускулатуру; они имеются также в вегетативной нервной системе. Безмякотные волокна у позвоночных животных принадлежат в основном симпатической нервной системе. Нервы обычно состоят как из мякотных, так и из безмякотных волокон, причем соотношение между числом тех и других в разных нервах различное. Например, вс многих кожных нервах преобладают безмякотные нервные волокна. Так, в нервах вегетативной нервной системы, например в блуждающем нерве, количество безмякотны> волокон достигает 80—95 %. Наоборот, в нервах, иннервирующих скелетные мышцы имеется лишь относительно небольшое количество безмякотных волокон. • На рис. 42 схематически показано строение миелинизированного нервного волокна Как видно, оно состоит из осевого цилиндра и покрывающей его миелиновой. оболочки Поверхность осевого цилиндра образована плазматической мембраной, а его содержимое представляет собой аксоплазму, пронизанную тончайшими (диаметром 10—40 нм) нейрофибриллами (и микротубулами), между которыми находится большое количестве митохондрий и микросом. Диаметр нервных волокон колеблется от 0,5 до 25 мкм. Как показали электронно-микроскопические исследования, миелиновая оболочкг создается в результате того, что миелоцит (шванновская клетка) многократно оберты вает осевой цилиндр (рис. 43, I), слои ее сливаются, образуя плотный жировой футляр — миелиновую оболочку. Миелиновая оболочка через промежутки равной длины прерыва ется, оставляя открытыми участки мембраны шириной примерно 1 мкм. Эти участм получили название перехватов (перехваты Ранвье). Длина межперехватных участков, покрытых миелиновой оболочкой, примерне пропорциональна диаметру волокна. Так, в нервных волокнах, имеющих диаметр 10— 20 мкм, длина промежутка между перехватами составляет 1—2 мм. В наиболее тонки; волокнах (диаметром 1—2 мкм) эти участки имеют длину около 0,2 мм. Безмякотные нервные волокна не имеют миелиновой .оболочки, они изолировань друг от друга только шванновскими клетками. В простейшем случае одиночный миелоцит окружает одно безмякотное волокно. Часто, однако, в складках миелоцита оказываете? несколько тонких безмякотных волокон (рис. 43, II). Миелин Перехват Ранвье 6Е Ядро Шванковской Шванновская нлетни оболочка Рис. 42. Строение миелинизированного нервного волокна (схема). 3 Физиология человека 'ис. 43. Роль миелоцита (шванновской клетки) в образовании миелиновой оболочки в мякотных .ервных волокнах. Показаны последовательные стадии спиралеобразного закручивания миелоцита округ аксона (I). Взаимное расположение миелоцитов и аксонов в безмякотных нервных волоках (II). ФИЗИОЛОГИЧЕСКАЯ РОЛЬ СТРУКТУРНЫХ ЭЛЕМЕНТОВ МИЕЛИНИЗИРОВАННОГО НЕРВНОГО ВОЛОКНА Можно считать доказанным, что в процессах возникновения и проведения нервного iMпульса основную роль играет поверхностная мембрана осевого цилиндра. Миелиновая (болонка выполняет двоякую функцию: функцию электрического изолятора и трофиче- :кую функцию. Изолирующие свойства миелиновой оболочки Связаны с тем, что миелин ;ак вещество Липидной природы препятствует прохождению ионов и потому обладает >чень высоким сопротивлением. Благодаря существованию миелиновой оболочки юзникновение возбуждения в мякотных нервных волокнах' возможно не на всем протя- кении осевого цилиндра, а только в ограниченных участках— перехватах узла (пережата Ранвье). Это имеет важное значение для распространения нервного импульса даоль волокна. . Трофическая функция; миелиновой оболочки, по-видимому, состоит в том, что она фннимает участие в процессах регуляции обмена веществ и роста осевого цилиндра. Nat * 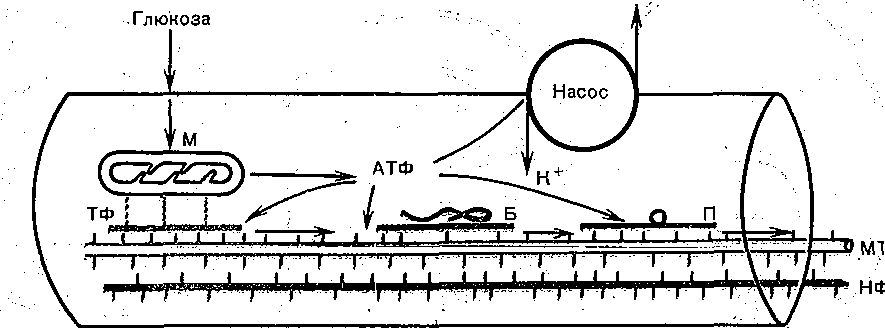 Рис. 44. Гипотетический транспортный механизм нервного волокна. Предполагается, что микротубулы (МТ) и нейрофиламенты (НФ) образованы миозином, а тонкие транспорт ныё филаменты — актином. При расщеплении АТФ транспортные филаменты скользят вдоль микротрубочек и таким образом переносят прикрепившиеся к ним митохондрии (М), молекулы белка (Б) или пузырьки (П] с медиатором. АТФ продуцируется митохондриями в результате распада глюкозы, проникающей в волокно Энергия АТФ частично используется также натриевым насосом поверхностной мембраны. Нейрофибриллы, микротубулы и транспортные филаменты обеспечивают транспорт различных веществ, и некоторых клеточных органелл по нервным волокнам от тела нейрона к нервным окончаниям и в обратном направлении. Так, по аксону из тела клетки на периферию транспортируются: белки, формирующие ионные каналы и насосы; возбуждающие и тормозные медиаторы; митохондрии- Подсчитано, что через поперечный разрез среднего по диаметру аксона в течение суток перемещается примерно 1000 митохондрий. Обнаружено, что нейрофибриллы образованы сократительным белком актином, а микротубулы — белком тубулином. Предполагают, что микротубулы, взаимодействуя с нейрофибриллами, выполняют в нервном волокне ту же роль, которую в мышечном волокне играет миозин. Транспортные филаменты, образованные актином, «скользят» вдоль микротубул со скростью 410 мкм/сут. Они связывают различные вещества (например, белковые молекулы) или клеточные органеллы (митохондрии) и переносят их вдоль волокна (рис. 44). Так же как и мышечный сократительный аппарат, транспортная система нервного волокна использует для своей работы энергию АТФ и нуждается в присутствии ионов Са2+.в цитоплазме. ПЕРЕРОЖДЕНИИ НЕРВНЫХ ВОЛОКОН ПОСЛЕ ПЕРЕРЕЗКИ НПРВЛ Нервные волокна не могут существовать вне связи с телом нервной клетки: перерезка нерва ведет к гибели тех волокон, которые оказались отделенными от тела клеток. У теплокровных животных уже через 2—3 сут после перерезки нерва периферический его отросток утрачивает способность к проведению нервных импульсов. Вслед за этим начинается дегенерация нервных волокон, причем миелиновая оболочка претерпевает жировое перерождение. Это выражается в том, что мякотная оболочка теряет миелин, который скапливается-в виде капель; распавшиеся волокна и их миелин рассасываются и на месте нервных волокон остаются тяжи, образованные леммоцитом (шванновской клеткой). Все эти изменения впервые были описаны английским врачом Валлером и названы по его имени валлеровским перерождением. Регенерация нерва происходит очень медленно. Леммоциты, оставшиеся на месте дегенерировавших нервных волокон, начинают разрастаться вблизи места перерезки по направлению к центральному отрезку нерва. Одновременно перерезанные концы аксонов центрального отрезка образуют так называемые колбы роста — утолщения, которые растут в направлении периферического отрезка. Часть этих веточек попадает в старое ложе перерезанного нерва и продолжает расти в этом ложе со скоростью 0,5—4,5 мм в сутки до тех пор, пока не дойдет до соответствующей периферической ткани или органа, где волокна образуют нервные окончания. С этого времени восстанавливается нормальная иннервация органа или ткани. В различных органах восстановление функции после перерезки нерва наступает в разные гроки. В мышцах первые признаки восстановления функций могут появиться через 5-—6 нед; окончательное восстановление происходит много позднее, иногда через год. • ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ При изучении проведения возбуждения по нерву было установлено несколько необходимых условий и правил (законов) протекания этого процесса. Анатомическая и физиологическая непрерывность волокна. Проведение импульсов возможно лишь при условии анатомической целостности волокна, поэтому как перерезка нервных волокон, так и любая травма поверхностной мембраны нарушают проводимость. Непроводимость наблюдается также при нарушении физиологической целостности волокна (блокада натриевых каналов возбудимой мембраны тетродотоксином или местными анестетиками, резкое охлаждение и т. п.). Проведение нарушается и при стойкой деполяризации мембраны нервного волокна ионами К, накапливающимися при ишемии в межклеточных щелях. Механическай травма, сдавливание нерва при воспали- гельном отеке тканей могут сопровождаться частичным или полным нарушением функции проведения. Двустороннее проведение. При раздражении нервного волокна возбуждение рас- фостраняется по нему и в центробежном, и в центростремительном направлениях. Это юказывается следующим опытом. К нервному волокну, двигательному или чувствительному, прикладывают две пары электродов, связанных с двумя электроизмерительными приборами А и Б (рис. 45). Раздражение наносят между этими электродами. В результате двустороннего проведе- > 1ия возбуждения приборы зарегистрируют прохождение импульса как под электродом А, гак и под электродом Б. Двустороннее проведение не является только лабораторным феноменом. В естественных условиях потенциал действия нервной клетки возникает в той ее части, где тело лереходит в ее отросток — аксон (так называемый начальный сегмент). Из начального :егмента потенциал действия распространяется двусторонне: в аксоне по направлению < нервным окончаниям и в тело клетки по направлению к ее дендритам. Изолированное проведение. В периферическом нерве импульсы распространяются ю каждому волокну изолированно, т. е. не переходя с одного волокна на другое и оказы- !ая действие только на те клетки, с которыми контактируют окончания данного нервного юлокна. Это имеет очень важное значение в связи с тем, что всякий периферический 1ервный ствол содержит большое число нервных волокон — двигательных, чувствитель- 1ых и вегетативных, которые иннервируют разные, иногда далеко отстоящие друг от фуга и разнородные по структуре и функциям клетки и ткани. Например, блуждающий jepB иннервирует все органы грудной полости и значительную часть органов брюшной юлости, седалищный нерв — всю мускулатуру, костный аппарат, сосуды и кожу нижней сонечности. Если бы возбуждение переходило внутри нервного ствола с одного волокна ia другое, то в" этом случае нормальное функционирование периферических органов I тканей было бы невозможно. Изолированное проведение в отдельных волокнах смешанного нерва может быть доказано фостым опытом на скелетной мышце, иннервированной смешанным нервом, в образовании кото- >ого участвует несколько спинномозговых корешков. Если раздражать один из этих корешков, :окращается не вся мышца, как это было бы в случае перехода возбуждения с одних нервных юлокон на другие, а только те группы мышечных волокон, которые иннервированы раздражаемым (орешком. Еще более строгое доказательство изолированного проведения возбуждения может быть юлучено при отведении потенциалов действия от различных нервных волокон нервного ствола; Изолированное проведение нервного импульса обусловлено тем, что сопротивление •кидкости, заполняющей межклеточные щели, значительно ниже сопротивления мем- Рис. 45. Схематическое изображение опыта для доказательства двустороннего проведения импульса в нерве. Объяснение в тексте. браны нервных волокон. Поэтому основная часть тока; возникающего между возбужден ным {деполяризованным) и покоящимися участками возбудимой мембраны, проходи по межклеточным щелям, не заходя в соседние волокна. Проведение возбуждения в немиелинизированных и миелинизированных нервных волокнах В безмякотных нервных волокнах возбуждение распространяется непрерывно вдол всей мембраны, от одного возбужденного участка к другому, расположенному phaoiv В отличие от этого в миелинизированных волокнах потенциал действия может распрс страняться только скачкообразно, «перепрыгивая» через участки волокна, покрыты изолирующей миелиновой оболочкой. Такое проведение называется сальтаторным. Прямые электрофизиологические исследования, проведенные Като (1924), а зате! Тасаки (1953) на одиночных миелинизированных нервных волокнах лягушки, пoкaзaл^ что потенциалы действия в этих волокнах возникают только в перехватах, а уча стки между перехватами, покрытые миелином, являются практически невозбудимыми Плотность натриевых каналов в перехватах' очень велика: на 1 мкм2 мембраш насчитывается около 10 000 натриевых каналов, что в 200 раз превышает плотност их в мембране гигантского аксона кальмара. Высокая плотность натриевых канало является важнейшим условием сальтаторного проведения возбуждения. Схема на рис. 4 позволяет понять, каким образом происходит «перепрыгивание» нервного импульса одного перехвата на другой. ^ В состоянии покоя наружная поверхность возбудимой мембраны всех перехвато] (перехватов Ранвье) заряжена положительно. Разности потенциалов между сс седними перехватами не существует. В момент возбуждения поверхность мембран! перехвата А становится заряженной электроотрицательно по отношению к поверхноел мембраны соседнего перехвата Б. Это приводит к возникновению местного (локального электрического тока, который идет через окружающую волокно межтканевую жидкость мембрану и аксоплазму в направлении, показанном на рис. 46 стрелкой. Выходящи! через перехват Б ток возбуждает его, вызывая перезарядку мембраны. В перехвате I возбуждение еще продолжается, и он на время становится рефрактерным. Поэтом; перехват Б способен привести в состояние возбуждения только следующий пере хват В и т. д. «Перепрыгивание» потенциала действия через межперехватный участок оказыва ется возможным только потому, что амплитуда потенциала действия в каждом перехват в 5—6 раз превышает пороговую величину, необходимую для возбуждения соседней перехвата. При определенных условиях потенциал действия может «перепрыгнуть: не только через один, но и через два межперехватных участка. Такое наблюдается в частности, в том случае, если возбудимость соседнего перехвата снижена каким-либ' фармакологическим агентом, например новокаином, кокаином и др. А Б 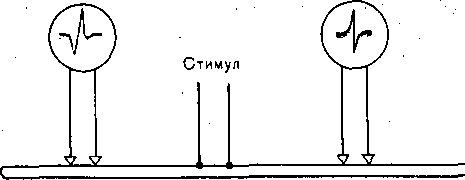 Время, необходимое для передачи возбуждения от одного перехвата другому примерно /одинаковое у волокон различного диаметра, (при температуре 24 °С он< составляет около 0,07 мс). Длина межперехватных участков, как отмечалось, пропор циональна диаметру нервного волокна. Отсюда следует, что в миелинизированны Рис. 46. Сальтаторное распространение возбуждения в мякотном нервном волокне от перехвата к перехвату. Стрелками показано направление тока, возникающего между возбужденным (А) и соседним покоящимся (Б) перехватом. волокнах скорость проведения нервного импульса примерно пропорциональна их диаметру. В этом отношении миелинизи- рованные волокна 'отличаются от безмякотных, у которых скорость проведения пропорциональна не диаметру, а корню квадратному из его величины.  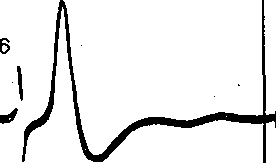 ишшшш! ис. 47. Ответы седалищ- jro нерва лягушки на шктрические стимулы врастающей силы. тведение потенциалов дей- •вия двухфазное. В начале 1 ждой записи виден арте- акт раздражения. Сила то- i увеличивается постепенно г 1 к 6. Отметка времени — мс. Проведение возбуждения по миелинизированному нервному волокну часто сравнивают с передачей сигналов по электрическому кабелю с ретранслирующими генераторами '(например, трансатлантическому кабелю). Действительно, участки нервного волокна между перехватами по своим электрическим свойствам подобны кабелю, погруженному в жидкость, обладающую высокой электропроводностью. Внутренним проводником является аксоплазма, внешним — межклеточная жидкость, а изолятором — жировая миелиновая оболочка. Импульс, проходящий между перехватами, представляет собой импульс электрического тока. Перехваты Ранвье играют роль ретранслирующих-генераторов, т. е. промежуточных усилительных станций линии связи. При передаче сигнала каждый следующий перехват возбуждается импульсом, генерируемым предыдущим, вырабатывает новый импульс и передает его .по волокну. Поскольку сопротивление.внутреннего проводника на единицу длины очень, велико (в 106 раз больше, чем медной проволоки того диаметра), ретранслирующие генераторы должны быть расположены близко друг к другу, иначе импульс угаснет. Предположение о скачкообразном распространении возбуждения в нервных волокнах впервые было высказано Б. Ф. Вериго (1899). Такой способ проведения имеет ряд преимуществ по сравнению с непрерывным проведением в безмякотных волокнах: во-первых, «перепрыгивая» через сравнительно большие участки волокна, возбуждение может распространяться со значительно большой скоростью, чем при непрерывном проведении: по безмякотному волокну того же диаметра; во-вторых скачкообразное распространение является энергетически более экономным, поскольку в состояние активности приходит не вся мембрана, а только ее небольшие -участки в области перехватов, имеющие ширину менее 1 мкм. Потери ионов (в расчете на единицу длины волокна), сопровождающие возникновение потенциала действия в таких ограниченных участках мембраны, очень невелики, а следовательно, малы и энергетические затраты на работу натрий-калиевого насоса, необходимые для восстановления измененных ионных соотношений между внутренним содержимым нервного волокна и тканевой жидкостью.. • СОСТАВНОЙ ХАРАКТЕР ПОТЕНЦИАЛА ДЕЙСТВИЯ НЕРВНОГО СТВОЛА И КЛАССИФИКАЦИИ НЕРВНЫХ ВОЛОКОН Амплитуда электрических импульсов, отводимых от 'целого нервного ствола, ависит от силы приложенного раздражителя. Раздражителю слабой силы соответствует 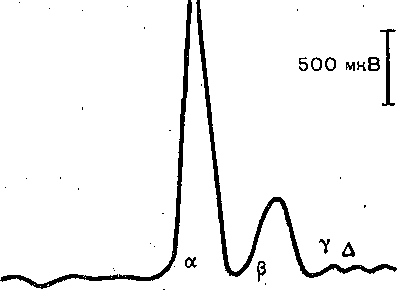 Рис. 48. Комплекс составного потенциала действия смешанного нервного ствола лягушки (по Эрлангеру и Гассеру). 175 150 125 100 75 50 25 О bhj > ' ' ■ ' 25 50 75 100 125 150 175 200 225 250 275 300 Рис. 49. Схематическоое изображение всех сс ставных частей потенциала действия смешаь ного нерва (Объяснение в тексте). небольшой ответ; по мере усиления раздражения, амплитуда потенциала возрастает, достигая максимальной величины, и затем остается постоянной, несмотря на дальнейшее увеличение силы раздражителя (рис. 47). Объясняется это тем, что электрический ответ целого нервного ствола является алгебраической суммой потенциалов действия отдельных его волокон. В каждом волокне амплитуда потенциала действия не зависит от силы раздражения в соответствии с законом «все йли ничего». Пороги раздражения отдельных волокон отличаются друг от друга. При слабой силе стимула возбуждение возникает в наиболее возбудимых, поверхностно расположенных нервных волокнах. Усиление стимула приводит к увеличению чис^а возбужденных волокон, поэтому суммарный ответ на раздражение увеличивается до тех пор, пока все волокна не вовлекаются в реакцию Такую картину можно наблюдать, если отводящие, электроды расположить на нерве вблизи раздражающих электродов. При увеличении расстояния между этими двумя парами электродов суммарный потенциал действия начинает расчленяться на несколько отдельных колебаний, которые становятся наиболее отчетливо выраженными при удалении отводящих электродов на 10—15 см от места раздражения (рис. 48). Впервые это явление было подробно изучено Эрлангером и Гассером (1937). Они показали, что причиной расчленения суммарного потенциала действия на компоненты является неодинаковая скорость проведения.возбуждения по разным волокнам, вследствие чего к отводящим электродам нервные импульсы поступают по этим волокнам неодновременно. В настоящее время нервные волокна по скорости проведения возбуждения, длительности различных фаз потенциала действия и строению принято подразделять на три основных типа, обозначаемых буквами А, В и. С (рис. 49). Волокна типа А делятся на четыре подгруппы: а, р, у, б. Они покрыты миелиновой оболочкой. Наиболее толстые их них а-волокна (Аа), у теплокровных животных и человека они имеют диаметр 12—22 мкм и характеризуются значительной скоростью проведения, возбуждения — 70—120 м/с. Такие волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от определенных рецепторов мышц к соответствующим нервным центрам. Пик потенциала действия волокна Аа у теплокровных длится 0,4—0/5 мс. После егс окончания развивается следовая деполяризация, которая продолжается 15—20 мс и переходит в следовую гиперполяризацию длительностью около 40—60 мс. Три другие группы волокон типа А: Ар, Ау и А6 — имеют меньший диаметр, меньшую скорость проведения и более длительный потенциал действия. Это преимущественнс чувствительные волокна, проводящие возбуждение от различных рецепторов (тактильных, некоторых болевых, температурных и рецепторов внутренних органов) в ЦНС. 7,1 Исключение составляют лишь у-волокна, значительная часть которых проводит юзбуждение в центробежном направлении от клеток спинного мозга к так называемым !нтрафузальным мышечным волокнам, входящим в состав рецепторов мышц — мышеч- 1ых веретен (см. рис. 51, А). К волокнам типа В относятся миелинизированные, преимущественно преганглио- [арные, волокна вегетативной нервной системы. Скорость проведения возбуждения ; этих волокнах у теплокровных животных составляет 3—18 м/с. Продолжительность ютенциала действия волокон типа В (1—2 мс) примерно в 3 раза превышает длитель- юсть потенциала действия волокон типа А. Отличительной особенностью этих волокон [вляется то, что в них не обнаруживается фаза следовой деполяризации: нисходящее олено пика непосредственно переходит в следовую гиперполяризацию, которая в ряде лучаев продолжается более 100 мс. К волокнам типа С относят безмякотные нервные волокна очень малого диаметра примерно 1 мкм). Скорость проведения возбуждения в этих волокнах не более 3 м/с. юльшинство волокон типа С — это постганглионарные волокна симпатической нервной истемы. К волокнам типа С относят также те нервные волокна, которые участвуют в прове- ,ении возбуждения от болевых рецепторов и некоторых рецепторов холода, тепла давления. Потенциалы действия волокон этого типа характеризуются наибольшей продолжи- ельностью (2 мс у теплокровных животных). Они имеют длительную фазу следовой .еполяризации (50—80 мс), сопровождающуюся еще более продолжительной (300 — 000 мс) следовой гиперполяризацией. Все эти данные суммированы в табл. 3. ТаблицаЗ Свойства различных нервных волокон теплокровных
ИССЛЕДОВАНИЕ СКОРОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВНЫМ ВОЛОКНАМ У ЧЕЛОВЕКА Скорость проведения возбуждения по нервным волокнам может быть определена у человек; сравнительно несложным путем. Для определения скорости проведения по двигательным волокнал используется электрическая стимуляция нерва через кожу в тех местах, где он расположен неглу боко. Используя электромиографическую методику, записывают электрический ответ мышцы н; это раздражение. Латентный период ответа в основном зависит от скорости проведения по нерву Измерив его, а также расстояние между стимулирующими и отводящими электродами, можн< рассчитать скорость проведения. -Более точно ее можно определить по разности латентного ответ; при раздражении нерва в двух точках. Для определения скорости.проведения по чувствительны!* волокнам наносят кожное электрическое раздражение, а ответ отводится от нерва. В связи с тем что высокая скорость проведения по нервным волокнам обусловлена миелиновой оболочкой, нарушения ее, наблюдающиеся при ряде заболеваний нервной системы, сопровождают^ снижением скорости проведения возбуждения. Поэтому методика определения скорости проведена по нервным волокнам широко используется в неврологических клиниках для диагностически; целей. 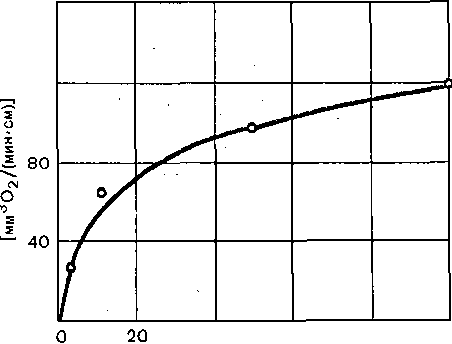 О 120 40 60 Частота стимуляции Рис. 50. Повышение потребления кислорода нервом в зависимости от частоты стимуляции. ХИМИЧЕСКИЕ ИЗМЕНЕНИЯ В НЕРВЕ В ПОКОЕ И ПРИ ПРОВЕДЕНИИ ВОЗБУЖДЕНИЯ Нерв в состоянии покоя потребляет кислород и выделяет углекислоту. Если один .участок нерва поместить в атмосферу азота, а другой оставить в воздухе, то уж< через несколько минут можно обнаружить, что поверхность нерва, лишенная кислорода приобретает электроотрицательный заряд. Объясняется это тем, что в бескислородной среде вследствие изменения обменных процессов нарушается работа натрий-калиевогс насоса, в результате чего происходит прогрессирующая Деполяризация мембраны нерв ных волокон. Примерно через 11 /2 ч потенциал покоя падает настолько, что проведе ние нервных импульсов на этом участке нерва полностью прекращается. При возвра щении нерва в атмосферу кислорода потенциал покоя быстро восстанавливается и даж< на некоторое время становится больше исходной величины. Одновременно восстанавли вается и проведение возбуждения. Возбуждение нерва сопровождается повышением потребления кислорода, при чем по мере увеличения частоты раздражения поглощение кислорода возрастав' (рис. 50). Вместе с тем отмечаются увеличение распада богатых энергией фосфор ных соединений — аденозинтрифосфата и креатинфосфата и повышенное обра зование молочной кислоты (за счет анаэробного расщепления глюкозы и гликогена) При возбуждении усиливается также белковый обмен в нервных волокнах, о чем свиде-, тельствует, в частности, выделение значительных количеств аммиака. Предполагают, что аммиак образуется в результате расщепления глутамина. Интенсивное раздражение нерва вызывает, кроме того, усиление обмена нуклеиновых кислот и фосфолипидов. При раздражении большинства мякотных нервных волокон из них высвобождается аце- тилхолин, из безмякотных симпатических нервных волокон —- норадреналин. Наличие ацетилхолина в одних" нервных волокнах и норадреналина в других свидетельствует о химической гетерогенности разных типов нервных волокон. В нервных окончаниях ацетилхолин и норадреналин являются химическими передатчиками нервного импульса — медиаторами.. Синтезируемые в соме нервных клеток медиаторы диффундируют вдоль этих волокон к нервным окончаниям, из которых они и выделяются при возбуждении. ТЕПЛОПРОДУКЦИЯ НЕРВА О динамике обмена веществ нерва в покое и при возбуждении можно, судить по его теплопродукции, которую впервые удалось зарегистрировать Хиллу в 1926 г. при помощи специально сконструированных высокочувствительных термоэлементов. Теплообразование седалищного нерва лягушки, измеренное в покое, составляет 4,14-10_3 кал на 1 г нерва .в минуту при 20 °С. Теплообразование безмякотного нерва краба несколько выше: 1 • Ю-2 кал на 1 г в минуту при 20 °С. Это согласуется с данными о значительно большем потреблении кислорода безмякотными нервами'в покое. Если поместить, нерв в атмосферу азота, теплообразование покоя резко снижается. ■ - При раздражении нерва теплообразование значительно возрастает. Так же как и в мышце< тепло выделяется в две фазы, которые обозначаются как начальное и запаздывающее теплообразование. ' Новейшие исследования, проведенные при помощи высокочувствительной и малоинерционной аппаратуры, показали, что начальное теплообразование непосредственно связано с процессом генерации потенциала действия. Так установлено, что подъем потенциала действия .сопровождается выделением небольшой порции тепла, а окончание пика — его поглощением. Запаздывающее теплообразование после ритмического раздражения нерва продолжается десятки минут. УТОМЛЕНИЕ НЕРВА Впервые Н. Е.; Введенский установил, что нерв в атмосфере воздуха сохраняет способность к проведению возбуждений даже при-многочасовом (около 8 ч) непрерывном раздражении. Это свидетельствует о том, что нерв в атмосфере воздуха практически не утомляем или. малоутомляем. Относительная, неутомляемость нерва отчасти зависит от того, что он тратит при своем возбуждении сравнительно мало энергии. Так, I г нерва лягушки выделяет при максимальном раздражении только на 20—100 % больше тепла, чем в покое. Такой прирост очень незначителен по сравнению с наблюдаемым при возбуждении мышцы. Благодаря этому процессы ресинтеза в нерве в состоянии покрывать его относительно малые расходы энергии при возбуждении даже в том случае, если оно длится много часов. , Энергетические траты при возбуждении нервных волокон связаны главным образом с работой натрий-калиевого насоса, который активируется поступлением внутрь цитоплазмы Na+. В условиях нормального кровоснабжения нерва натрий-калиевый насос обеспечивает устойчивое поддержание ионного состава цитоплазмы, так как число ионов N-a+, поступающих внутрь волокна, и К+, покидающих волокно при Каждом импульсе, очень мало по сравнению с общим их содержанием в цитоплазме и межклеточной жидкости. Если принять, что число ионов, пересекающих единицу площади мембраны, в различных волокнах одинаково, то в этом случае изменение концентрации этих ионов в цитоплазме должно быть обратно пропорционально диаметру волокна. Поэтому волокно диаметром 0,5 мкм при каждом импульсе должно терять 1/юоо содержания К+ вместо '/i оо'о ооо, как это наблюдается в гигантских аксонах кальмара. Этим, по-видимому, и объясняется тот факт, что тонкие нервные волокна .утомляются значительно быстрее, чем толстые. НЕРВНО-МЫШЕЧНАЯ ПЕРЕДАЧА В предыдущих разделах было показано, что проведение возбуждения в нервных и мышечных волокнах осуществляется при помощи электрических импульсов, распространяющихся по поверхностной мембране. Передача возбуждения с нервного волокна 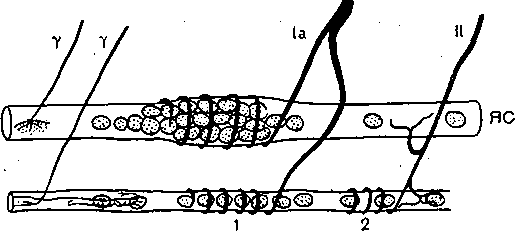 Рис. 51. Иннервация интрафузальных мышечных волокон веретена млекопитающих и ответ перви^ ного окончания веретена человека на изометрическое сокращение мышцы с нагрузкой. А: ЯС и ЯЦ — ядерно-сумчатые и ядерно-цепочечные волокна, 1,2—первичные и вторичные окончани* 1а и Н-афферентные нервные волокна, 7-афференты. Б: а — импульсация одиночного афферентного волокн 1а; б —• суставной угол (сгибание-пальца); в — электромиограмма сгибателя пальца; г — отметка времен 1 с. Калибровка: 100 мкВ, 10°, 0,2 мВ. на мышечное основана на совершенно ином механизме. Она происходит в результат выделения нервными окончаниями- химических соединений—медиаторов (передатчи ков) нервного импульса. У человека, как и у всех позвоночных, роль медиатора в скелет ных мышцах играет ацетилхолин. - Предположение, - что в передаче возбуждения в нервно-мышечном соединени принимают участие какие-то химические агенты, впервые было высказано А. Ф. Само? ловым в 1924 г. Позднее Дейл :(1936), показал, что при раздражении двигательног нерва в его окончаниях в скелетной мышце происходит выделение ацетилхолина. Наконе! было установлено, что ацетилхолин, подведенный к области нервно-мышечного соед& нения,, деполяризует мембрану мышечного волокна и при достаточно высокой концеь трации вызывает распространяющееся возбуждение и сокращение мышцы. , НЕРВНО-МЫШЕЧНОЕ СОЕДИНЕНИЕ (СИНАПС) Структурное образование, обеспечивающее переход возбуждения с нервного вс локна на иннервируемую им клетку —мышечную, нервную или железистую, получил название синапса. 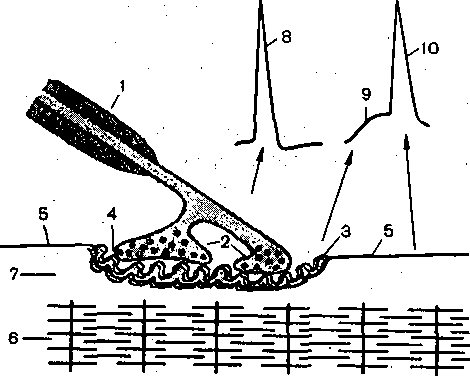 Электронно-микроскопические исследования выявили, что так же как в ЦНС, н периферии синапсы состоят из трех.основных элементов: пресинаптической мембрань, постсинапттеской мембраны и синаптической -щели (рис. 52). • Рис. 52. Взаимоотношения между нервным волокном, нервным окончанием и скелетным мышечным волокном (схема). . \  Ш) ЯЦ В I — миелинизированное нервное волокно; 2 — нервное окончание с пузырьками медиатора; 3 — постсинаптическая мембрана мышечного волокна; 4 — синаптическая щель;. 5 — внесинаптическая мембрана мышечного волокна; 6— миофибриллы; 7 — саркоплазма; 8 — потенциал действия нервного волокна; 9 — потенциал концевой пластинки (постсина'птический потенциал); 10 —потенциал действия мышечного волокна. Пресинаптической называется мембрана, покрывающая нервное окончание, которое лредставляет собой своеобразный нейросекреторный аппарат. Здесь содержится и зыделяется медиатор, оказывающий возбуждающее или тормозящее действие на яннервируемую клетку. \ " В скелетной мышце позвоночных двигательное м.иелиновоёНервное волокно разветвляется веерообразно на концевые безмякотные волокна диаметром около 1,5 мкм. На всем этом концевом участке нервное волокно (пр.есинаптическое окончание) образует :инаптическое соединение. с мышечным волокном. Вся область мышечного волокна, з которой расположены синапсы, образованные двигательным нервным волокном, зазывают концевой пластинкой (бляшкой; пуговкой). В пресинаптических окончаниях медиатор ацетилхолин содержится в «пузырьках» диаметром около 50,нм..При достижении распространяющегося потенциала действия области пресинаптического окончания ацетилхолин освобождается из «пузырьков» и зыходит в синаптйческую щель. В механизме этого нейросекреторного процесса важную золь играют ионы Са2+: они поступают внутрь окончания из внеклеточной жидкости ю электровозбудимым кальциевым каналам, активируемым при деполяризации преси- заптической мембраны приходящим потенциалом действия. При этом наблюдается следующая цепь процессов: деполяризация пресинаптической мембраны при проведении нервного импульса — >- открывание кальциевых каналов —:—> вхождение ионов Са.2+ внутрь окончания —-—выделение медиатора в синаптическую щель. Ширина юследней примерно 50 нм; она заполнена межклеточной жидкостью, которая по составу триближается к плазме крови. Медиатор быстро диффундирует через щель, воздействуя на мембрану иннервируемого мышечного волокна. Та часть мембраны этого волокна, которая непосредственно граничит с нервным окончанием, называется постсинаптической. От мембраны, покрывающей остальную часть мышечного волокна, постсинапти- 1еская мембрана отличается тем, что не содержит электрически возбудимых ионных каналов и потому не . способна к генерации потенциала действия. Постсинаптическая мембрана обладает, однако, химической возбудимостью: на действие ацетилхолина она отвечает местным изменением проницаемости для ионов Na+ и К+, что приводит к развитию так называемого потенциала концевой пластинки (ПК,П). По своей природе эн аналогичен возбуждающим постсинаптическим Потенциалам, возникающим при передаче, возбуждения с одной нервной клетки на другую. ПКП порождает генерацию потенциала действия в мышечном волокне. Наличие химического звена в механизме нервно-мышечной передачи делает понятным два общих свойства синапсов: 1) возбуждение проводится через синапс только в одном направлении — с нерва на мышцу; 2) возбуждение проводится через синапс значительно медленнеечем по нервному волокну. Односторонность; проведения обусловлена наличием относительно широкой синап- гической щели, препятствующей проведению импульса с помощью локальных токов 2 мышцы на нерв. Замедление проведения через синапс объясняется тем, что это проведение является многоэтапным процессом: время затрачивается на секрецию медиатора, гго диффузию к постсинаптической мембране, активацию последней, рост ПКП до пороговой величины. |