• В механизме освобождения ионов Са
2* из ретикулума при возбуждении особую роль играет система
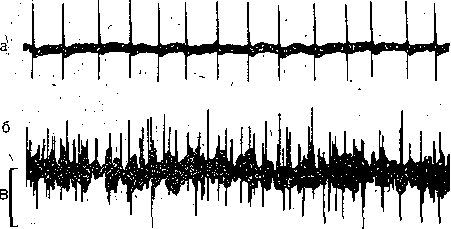
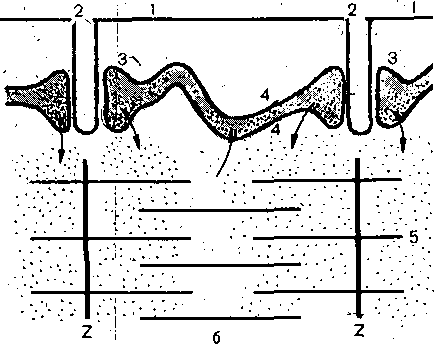
поперечных трубочек (Т-система), представляющих собой впячица'ния поверхностной. мембраны. Диаметр каждой трубочки: около. 0,05; мкм. На рис. 35 приведена , cxeiyia продольного среза через быстрое мы-, щечное волокно лягушки. Видно, что по обе стороны от поперечной трубочки расположены боковые (терминальные)
цистерны ретикулума: Вместе с трубочкой они образуют так называемые триады. Мембрана поперечных трубочек по своим свойствам сходна с поверхностной мембраной; она содержит электровозбудимые натриевые каналы и способна к генерации и проведению потенциала действия. Во время возбуж-
юния потенциал действия с поверхностной мембраны распространяется вдоль мембраны юперечных трубочек в глубь волокна и при помощи особого, пока еще полностью не изу- 4енного, механизма вызывает освобождение ионов Са
2+ из боковых цистерн. Боковые хистерны расположены таким образом, что освободившиеся Ионы Са
2+ попадают непо-> :редственно в ту область, где происходит образование актомиозина. '
Как отмечалось, начало мышечного сокращения приурочено к первой трети восходя- дего колена потенциала действия, а именно к моменту, когда внутренний потенциал юлокна возрастает с исходных —90 мВ до примерно —50 мВ. Этот потенциал является юроговым для возникновения механического ответа. Предполагают, что именно при дестижении указанного уровня деполяризации концентрация свободных ионов Са
2+ в
; миофибрилле достигает критической величины, необходимой для начала взаимодействия 1Ктиновых и миозинЬвых нитей.
v '
' Процесс Освобождения Са
2+ прекращается после окончания пика потенциала дей-. ггвия. Тем не^менее сокращение продолжает нарастать до тех пор, пока активация каль- ^евого насоса ретикулума не вызовет снижения концентрации ионов Са
2+ в миоплазме. Гогда сокращение сменяется расслаблением. _
1Таким образом, последовательность событий, ведущих к сокращению, а затем расслаблению мышечного волокна/представляется в следующем виде: раздражение —>- воз- адкновение потенциала действия -^ проведение его вдоль клеточной мембраны и в глубь юлокна по трубочкам-^-освобождение Са
2+ из боковых цистерн саркоплазматического )етикулума и диффузия его к'миофибриллам-vвзаимодействие («скольжение») актино- зых и миозиновых нитей, приводящее к укорочению миофибриллы активация кальциевого насоса-V снижение концентрации свободных ионов Са
2+ в саркоплазме -»- рас- :лабление миофибрилл. - , • \
Роль АТф в механизмах мышечного сокращенияВ процессе взаимодействия миозиновых И' актиновых ■ нитей в присутствии Са
2+ $ажную роль играет богатое энергией соединение — АТФ. Впервые В. А. Энгельгард \ М. Н. Любимова в 1939 г. нашли, что мышечный.белок — миозин — обладает свойствами фермента аденозйнтрифосфатазы. (АТФ-аза). Эти авторы также обнаружили, что юд влиянием АТФ изменяются и механические свойства миозина — резко увеличивается )астяжимосгь его нитей, В последующие годы был открыт белок актин и показано, что )Н активирует АТФ-.азную активность миозина.
В настоящее время взаимодействие АТФ с сократительными белками и ионами Са
2+ федставляют следующим образом. В покоящейся мышце; т. е. в условиях, когда концентрация ионов Са
2+ в миоплазме очень низкая, взаимодействию миозиновых и актиновых 1итей препятствуют молекулы белка тропонина, расположенные на актиновых нитях. Гропонин обладает очень высоким сродством к ионам Са^
+. Как Только концентрация За
2+ в миофибриллах повышается, тропонин связывает Са
2+ и изменяет свое расположе- ше на актиновой нити таким образом, что делает возможным ее взаимодействие с миози- ювой нитью. Формирующиеся при этом поперечные мостики перемещают актиновую шть лишь на 1 % ее длины. Чтобы обеспечить дальнейшее продвижение
нити и соответственно сокращение волокца, необходимо, чтобы эти мостики разъединились и прикре- 1ились к новым участкам актиновой нити. Такое разъединение мостиков осуществляется три расщеплении .молекул АТФ АТФ-а?ой миозина. Таким образом, взаимодействие лиозина и актина, возможное'при связывании Са
2+, тропонином, активирует АТФ-азу миозина, последняя расщепляет АТФ, а это приводит к разъединению миозина и актина.
В присутствии Са
2+ и;АТФ в миоплазме этот процесс многократно повторяется:'мостики ювторно образуются и расходятся, в результате чего актиновые нити «скользят» и мыше- шое волокно укорачивается.
Подсчитано, что при изотоническом сокращении скелетной мышцы лягушки поперечные мостики должны совершить за 0,1 с 50 таких движений, чт,обы длина каждого
саркомера волокна укоротилась на 50%. При каждом движении мостиков происходит ■ расщепление молекул АТФ. .
Таким образом, присутствие АТФ в мышце является обязательным условием дл* обратимости.связывания актина и миозина. Трупное окоченение возникает при услойии если концентрация АТФ в мышце падает ниже некоторой критической величины. Тогдг комплекс актин— миозин становится стабильным. ■
> Итак, энергия АТФ используется во, время деятельности скелетной мышцы для тре> основных процессов: 1)
работы натрий-калиевого насоса, обеспечивающего поддержание постоянства градиента концентрации ионов Na+ и К^ по обе стороны мембраны; 2)
процесса «скольжения» актиновых и миозиновых нитей, ведущёго
хк укорочению миофибрилл; 3)
работы кальциевого насо,са/необходимого для расслабления волокна. В.соответствии с.этим ферменты, расщепляющие АТФ, локализованы в трех различных структурах мышечного волокна: клеточной мембране, миозиновых нитях.и мембранах саркоплазматического ретикулума. , ' .
Ресинтез АТФ, непрерывно расщепляющейся в '.процессе деятельности мышцы осуществляется двумя основными путями. Первый состоит в ферментативном перенос* фосфатной группы от богатого энергией фосфорного соединения
креатинфо(;фата нг аденозиндифосфорную кислоту. Креатинфосфат содержится в-мышце в- значительно больших количествах, чем АТФ, и обеспечивает ее ресинтез в течение тысяч
долеР. секунды.^.Однако при интенсивной работе мышцы запасы креатинфосфата быстро истощаются, поэтому важен второй путь — более медленный
ресинтез АТФ. Он связан с Рликолитическими и окислительными процессами, протекающими в мышце как в условиях покоя, так и особенно интенсивно во время деятельности. Окисление
молочной и
пир о- виноградной кислот, образующихся в мышце во время ее сокращения, сопровождается фосфорилированием аденозиндифосфорной кислоты и креатина, т. е. ресинтезом креатинфосфата и АТФ. - '
Нарушение ресцнтеза АТФ ядами, подавляющими гликолитические и. Окислительные процессы,
ведет к полному исчезновению АТФ и креатинфосфата, вследствие чего кальциевый насос перестает работать. Концентрация Са
г+ в.области миофибрилл значительно
1 возрастает и мышца приходит в состояние длительного необратимого укорочения.
Теплообразование при сократительном процессе и энергия сокращенияОбразование тепла в мышечной ткани при работе было открыто Гельмгольцем и В. Я- Данилевским во второй половине, XIX в. В дальнейшем Хиллу с сотр
1- .удалось создать высокочувствительные приборы, которые позволили зарегистрировать и Измерить теплопродукцию мышц и нервов в покое и при возбуждении.
vПо рвоему происхождению и времени развития теплообразование было подразделено Хилдом на две основные фазы.
Первая фаза приблизительно в 1000 раз короче второй и называется фазой начального теплообразования. Она начинается с момента возбуждения мышцы ц продолжается в течение всего сокращения, включая фазу расслабления.
В. свою очередь начальное теплообразование может быть разделено на несколько частей: а) тепло активации; б) тепло укорочения; в) тепло расслабления.'
Тепло активации освобождается непосредственно после нанесения раздражения, но до сколько-нибудь^различимого сокращения мышечных волокон. Поэтому указанная порция тепла рассматривается как тепловой эффект тех химических процессов, которые переводят мышцу из невозбужденного состояния в активное!'При тетаническом сокращении тепло активации выделяется в течение всего времени раздражения мьщщы при каждом потенциале действия. Л, • ■
Тепло укорочения обусловлено самим сократительным процессом. Если путем силь-'
чного растяжения iv/ышцы воспрепятствовать ее сокращению, эта порция тепла не выде- '' ляе'гся.
V Тепло расслабления'связано с освобождением энергии в результате расслабления мышцы. Если мышца подняла груз во время сокращения, то по'окончании его количество выделяемого тепла увеличивается.
Вторая фаза теплопродукции длится несколько минут после расслабления и носит, название запаздывающего, или восстановительного, теплообразования. *
Юна связана С химическими, процессами, обеспечивающими.ресинтез АТФ. В опытах на мышцах, сокращающихся-в отсутствие кислорода, -Хилл показал, что в отличие от начального теплообразования, для которого кислород не нужен, 90 % восстановительного гепла образуется в результате окислительных процессов и лишь 10 % этого тепла обусловлены ^анаэробными процессами обмена веществ. Тепло восстановления по; своей
величине примерно равно количеству тепла, выделяемого мышцей во время сокращения. Это соответствие становится понятным^ если учесть, что химические процессы, обусловливающие восстановительное теплообразование, направлены на ресинтез АТФ, являющийся основным непосредственным источником энергии мышечного сокращения. Главную роль в ресинтезе АТФ и восстановительном теплообразовании играют процессы гликолиза и окислительного фосфор ил и Јюва ни я. Отравление мышцы монойодуксусной кислотой, прекращающей гликолитическое образование молочной и пировиноградной кислот, почти полностью выключает запаздывающее теплообразование и ресинтез АТФ даже в присутствии кислорода.
11  Скачать 7.39 Mb.
Скачать 7.39 Mb.