Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
 Скачать 7.39 Mb. Скачать 7.39 Mb.
|

РОЛЬ ОБМЕНА ВЕЩЕСТВ В ГЕНЕЗЕ И ПОДДЕРЖАНИИ ПОТЕНЦИАЛА ПОКОЯ (НАТРИЕВЫЙ НАСОС МЕМБРАНЫ) Несмотря на то что потоки Na+ и К+ через мембрану в покое малы, разность концентраций этих ионов внутри клетки и вне ее должна была бы в конечном ито^е выровняться, если бы в клеточной мембране не существовало особого молекулярного устройства—«натриевого насоса», которое обеспечивает выведение («выкачивание») из цитоплазмы проникающих в нее Na+ и введение («нагнетание») в цитоплазму К+, Натриевый насос перемещает Na+ и К+ против их концентрационных градиентов., т. е. совершает определенную работу. Непосредственным источником энергии для этой, работы является богатое.энергией (макроэргическое) соединение —■аденозинтрифосфорная кислота (АТФ), являющаяся универсальным источником энергии живых клеток. Расщепление АТФ производится макромолекулами белка — ферментом аденозинтрифосфатазой (АТФ-азой), локализованной в поверхностной мембране клетки. Энергия, выделяющаяся при расщеплении одной молекулы АТФ, обеспечивает выведение из клетки трех ионов К'а'1" взамен да два иона К+, поступающих в клетку снаружи. . Торможение активности АТФ-азы, вызываемое некоторыми химическими соединениями (например, сердечным гликозидом уабаином), нарушает работу насоса, вследствие чего клетка теряет К+ и обогащается Na+. К такому же результату приводит торможение окислительных и гликолитических процессов в клетке, обеспечивающих синтез АТФ.,В эксперименте это достигается при помощи ядов, ингибирующих указанные процессы. В условиях, нарушения кровоснабжения тканей, ослабления процесса тканевого дыхания происходит угнетение работы электрогенного насоса и как следствие накопление К+ в межклеточных щелях и деполяризация мембраны. Роль АТФ в механизме активного транспорта Na+ прямо доказана в опытах на гигантских нервных волокнах кальмара. Выло установлено, что путем введения внутрь волокна АТФ можно временно восстановить работу натриевого насоса, нарушенную ингибитором дыхательных ферментов цианидом. \ Первоначально полагали, что натриевый насос электронейтрален, т. е. число обмениваемых ионов Na+ и К+ равно. В дальнейшем,выяснилось, что на каждые три иона Na+, выводимые из клетки, в клетку поступает только два иона К+. Это-означает, что насос электрогенен: он создает на мембране разность потенциалов, суммирующуюся с потенцйадом покоя. - Этот вклад натриевого насоса в нормальную величину потенциала покоя у различных клеток не одинаков: 'он, по--видимом у, незначителен в нервных волокнах кальмара, но существен для потенциала покоя (составляет около 25% от полной величины) в гигантских нейронах моллюсков, гладких мышцах. Таким,образом, в формировании потенциала покоя натриевый насос играет двоякую роль: -1) создает и поддерживает транемембранный градиент концентраций Na+ и К+; 2) генерирует разность потенциалов, суммирующуюся с потенциалом, создаваемым диффузией JK+ по концентрационному градиенту. ПОТЕНЦИАЛ ДЕЙСТВИЯ Потенциалом Действия называют быстрое колебание мембранного потенциала, возникающее при Возбуждении нервных, мышечных и некоторых других клеток. В его основе лежат изменения ионной проницаемости мембраны. Амплитудами характер временных изменений потенциала действия мало зависят от силы вызывающего его раздражителя, важночлишь, чтобы эТа сила была не меньше некоторой критической величины, которая называется порогом раздражения. Возникнув в месте раздражения, потенциал действия распространяется вдоль нервного или мышечного волокна, не изменяя своей амплитуды. Налич-ие порога и независимость амплитуды потенциала действия от силы вызвавшего, его стимула получили название закона «все или ничего». 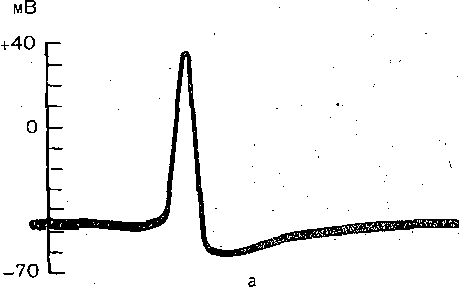 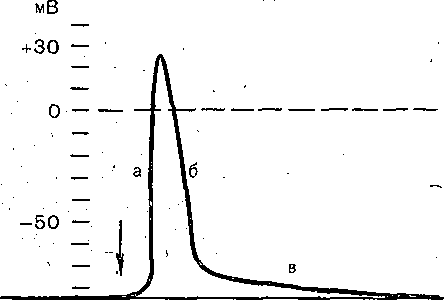 -100 —, Л Л П II И I Й 1 III I I I НЛ М А ЛЛ Рис. 3. Потенциал действия скелетного мышечного волокна, зарегистрированный с помощью внутриклеточного . микроэлектрода. а— фаза деполяризация, б — фаза рсполяризации, в — фаза следорой деполяризации (отрицательный следовой потенциал)\ Момент нанесения раздражения показан стрелкой. Рис. 4. Потенциал действия гигантского аксона кальмара . отводимый с помощью внутриклеточного электрода [Ходжкин А., 1965]. , , ■ - По вертикали отложены значения 'потенциала внутриклеточного электрода по отношению к его потенциалу в наружном растворе (в милливольтах); а — следовой положительный потенциал; б — отметка времени — 500 колебаний в 1 с. ' В естественных условиях потенциалы действия генерируются в нервных волокнах при раздражении рецепторов или возбуждении нервных клеток. Распространение потенциалов действия по нервным волокнам обеспечивает передачу информации в нервной системе. Достигнув нервных окончаний, потенциалы действия вызывают секрецию химических веществ (медиаторов) ^обеспечивающих передачу сигнала на мышечнце или нервные клетки. В мышечных клетках потенциалы действия инициируют цепь процессов, вызывающих сократительный акт. Ионы, проникающие в цитоплазму во время генерации потенциалов действия, оказывают регулирующее влияние на метаболизм клетки и, в частности, на'процессы синтеза белков, составляющих ионные каналы и ионные насосы. Для регистрации потенциалов действия используют вне- или внутриклеточные элект-. роды. При внеклеточном отведении электроды подводят к наружной поверхности волокна (клетки). Это позволяет обнаружить, что поверхность возбужденного участка на очень короткое время (в нервном волокне на тысячную долю секунды), становится заряженной отрицательно по отношению к соседнему покоящемуся участку. Использование внутриклеточных микроэлектродов позволяет количественно охарактеризовать изменения мембранного потенциала во время восходящей и нисходящей фаз потенциала действия. Установлено, что во время восходящей фазы (фаза деполяризации) происходит не просто исчезновение потенциала покоя (как это первоначально предполагали), а возникает разность потенциалов обратного знака: внутреннее содержимое клетки становится заряженным положительно по отношению к наружной, среде, иными, словами, происходит реёерсия' мембранного потенциала. Во время нисходящей фазы (фазы реполяризации) мембранный потенциал возвращается к своему исходному значению. На рис, 3 и 4 приведены примеры записей потенциалов действия в скелетном мышечном волокне лягушки и гигантском аксоне кальмара. Видно, что в момент достижения вершины (пика) мембранный потенциал составляет +30 + +40 мВ и пиковое колебание сопровождается длительными следовыми изменениями мембранного потенциала, после чего мембранный потенциал устанавливается на исходном уровне. Длительность пика потенциала действия у различных нервных и скелетных мышечных волокон варьи- рует от 0,5 до 3 мс, причем фаза реполяриза- ции продолжительнее фазы деполяризации. Длительность потенциала действия, особенно фазы реполяризации, находится в тесной зависимости от температуры: при ' охлаждении на 10 °С продолжительность пика увеличивается примерно в 3 раза. - Изменения мембранного потенциала, следующие за: пиком потенциала действия, называют следовыми потенциалами. ' х Различают два вида следовых потенциалов — следовую деполяризацию и следовую гиперполяризацию. Амплитуда следовых потенциалов обычно не превышает нескольких милливольт (5—10% от высоты пика), а длительность иХ1,у различных волокон составляет от нескольких миллисекунд до десятков и сотен секунд. " , Зависимость пика потенциала'действия и следовой деполяризации может быть рассмотрена на примере электрического ответа скелетного мышечного волокна. Из записи, приведенной на рис. 3, видно, что нисходящая фаза потенциала действия (фаза реполяризации) делится на две неравные части. Вначале падение потенциала происходит быстрота затем сильно замедляется. Этот медленный компонент нисходящей фазы потенциала действия называют следовой деполяризацией. ■ , . Пример следовой гиперполяризации мембраны, сопровождающей пик потенциала действия в одиночном (изолированном) гигантском нервном волокне кальмара, показан на рис. 4. В этом случае нисходящая фаза потенциала действия непосредственно переходит в фазу следовой гиперполяризации, амплитуда которой в данном случае достигает 15.мВ. Следовая гиперполяризация характерна для многих безмякотных нервных волокон холоднокровных и теплокровных животных. В миелинизированных нервных волокнах следовые потенциалы имеют более сложный характер. Следовая деполяризация может переходить в следовую гиперполяризацию, затем иногда .возникает новая деполяризация^ лишь после этого происходит полное восстановление потенциала покоя. Следовые потенциалы в значительно большей мере, чем пики потенциалов действия, чувствительны к изменениям исходного потенциала покоя, ионного состава среды, кислородного снабжения волокна и т. д. , Характерная особенность следовых потенциалов — их способность изменяться в процессе ритмической импульсации (рис. 5). - . . V '' - ' ' ИОННЫЙ МЕХАНИЗМ ВОЗНИКНОВЕНИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ В основе потенциала действия лежат последовательно развивающиеся'во времени изменения ионной проницаемости клеточной мембраны. Как отмечалось, в состоянии покоя проницаемость мембраны для калия превышает ее проницаемость для натрия. Вследствие этого поток К+ из цитоплазмы во внешний раствор превыщает противоположно направленный поток Na+. Поэтому наружная сторона мембраны в покое имеет положительный потенциал 'по Отношению к внутренней.  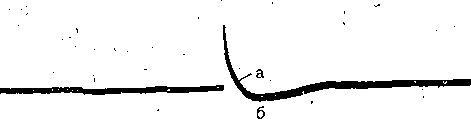 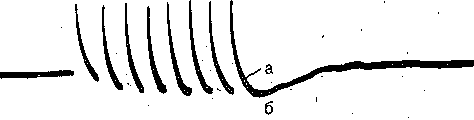 Рис; 5. Суммация следовых потенциалов в диаф- рагмальном нерве кошки при кратковременном его раздражении ритмическими импульсами.; Восходящая часть, потенциала действия не видна. Записи начинаются с отрицательных следовых потенциалов (а), переходящих в положительные потенциалы (б). Верхняя кривая —ответ на одиночное раздражение.. С увеличенем частоты стимуляции (от 10 до 250 в 1 с) следовой положительный потенциал (следовая гиперполяризация) резко возрастает. При действии .на клетку раздражителя проницаемость'мембраны для Na"1 резко повышается и в конечном итоге становится примерно в 20 раз больше проницаемости для К+- Поэтому поток Na+ из внешнего раствора в цитоплазму начинает превышать направленный наружу калиевый ток. Это приводит к изменению знака (реверсии) мембранного потенциала: внутреннее содержимое клетки становится заряженным положительно по отношению к ее наружной поверхности. Указанное изменение мембранного потенциала соответствует восходящей фазе потенциала действия (фаза деполяризации). Повышение проницаемости мембраны для Na+ продолжается лишь очень короткое время. Вслед за этим проницаемость мембраны для Na+ вновь понижается, а для К+ возрастает. \ Процесс, ведущий к понижению ранее увеличенной натриевой проницаемости мембраны, назван натриевой инактивацией. В результате инактивации поток Na+внутрь цитоплазмы резко ослабляется. Увеличение же калиевой проницаемо- :.■'.;.> сти вызывает усиление потОка К+ из цитоплазмы во внешний раствор. В итоге этих двух процессов и происходит реполяризация мембраны: внутреннее содержимое клетки вновь приобретает отрицательный заряд по отношению к наружному раствору. Этому изменению потенциала соответствует нисходящая фаза потенциала действия (фаза реполяри- зации). Одним из важных аргументов в пользу натриевой теории происхождения потенциалов действия был факт тесной зависимости1 его амплитуды от концентрации Na"1" во внешнем растворе. Опыты на гигантских нервных волокнах,, перфузируемых изнутри солевыми растворами, позволили получить прямое подтверждение правильности натриевой теории. Установлено, что при замене аксоплазмы солевым раствором, богатым К+> мембрана волокна не только удерживает нормальный потенциал покоя, но в течение длительного времени сохраняет способность генерировать сотни тысяч потенциалов действия нормальной амплитуды. Если "же К4" во внутриклеточном растворе частично заменить на Na+ и тем самым снизить градиент концентрации Na+ между наружной средой и внутренним раствором, амплитуда потенциала действия резко понижается. При полной замене К+ на Na+ волокно утрачивает способность генерировать потенциалы действия. \ Эти опыты не оставляют сомнения в том, что поверхностная мембрана действительно является местом возникновения потенциала как в покое, так и при возбуждении. Становится очевидным, что разность концентраций Na+ и К+ внутри и вне волокна является источником электродвижущей силы, обусловливающей возникновение потенциала покоя и потенциала действия. На рис. 6 показаны изменения натриевой и калиевой проницаемости мембраны во время генерации потенциала действия в гигантском аксоне кальмара. Аналогичные отношения имеют место в других нервных волокнах,-телах нервных клеток, а также в скелетных мышечных волокнах позвоночных животных. В скелетных мышцах ракообразных животных и гладких мышцах позвоночных в генезе восходящей фазы потенциала действия ведущую роль играют ионы Са2+. В клетках миокарда начальный подъем потенциала действия связан с повышением проницаемости мембраны для Na+, а плато потенциала действия обусловлено повышением проницаемости, мембраны и для ионов Са2+. О ПРИРОДЕ ИОННОЙ ПРОНИЦАЕМОСТИ МЕМБРАНЫ. ИОННЫЕ КАНАЛЫ 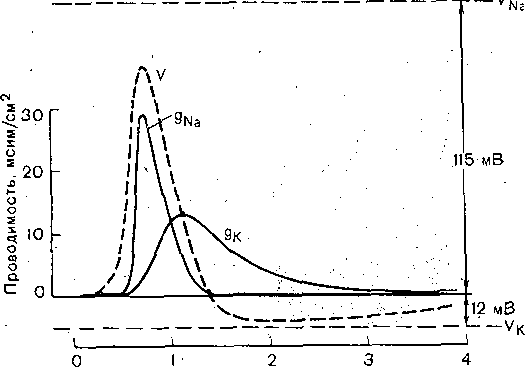 ■ _ Время, мс Рис. 6: Временной ход изменений натриевой (g^a) и калиевой (gk) проницаемости мембраны гигант- скогоаксона кальмара во время генерации потенциала действия (V). В основе рассмотренных изменений ионной проницаемости мембраны при генераций потенциала действия лежат процессы открывания и закрывания специализированных ионных каналов в мембране, обладающих двумя важнейшими свойствами: 1) избирательностью (селективностью) по отношению к определенным ионам; 2) электровозбуди мостью, т. е. способностью открываться и закрываться в ответ на изменения мембранного потенциала. Процесс открывания и закрывания канала имеет вероятностный характер (мембранный потенциал лишь определяет вероятность нахождения канала в открытом или закрытом состоянии). ' Так же как ионные насосы, ионные каналы образованы макромолекулами белков, пронизывающими липидный бислой мембраны. Химическая структура этих макромолекул еще на расшифрована, поэтому представления о функциональной организации каналов строятся пока главным образом косвенно — на основании анализа данных, полученных при исследованиях электрических явлений в мембранах и влияния на каналы различных химических агентов (токсинов, ферментов, лекарственных веществ и т. д.). Принято считать, что ионный канал состоит из собственно транспортной системы и так называемого воротного механизма («ворот»), .управляемого электрическим полем мембраны. «Ворота» могут находиться в двух положениях: они полностью закрыты илшпол- ностыо открыты, поэтому проводимость одиночного открытого канала—постоянная, величина, Суммарная проводимость мембраны для того или иного иона определяется числом одновременно открытых каналов, проницаемых для данного иона. ■ Это положение может быть записано следующим образом: gr. ■ /V-«7, ' гдеgi — суммарная проницаемость мембраны для внутриклеточного иона; N ■—общее число соответствующих ионных каналов (в данном участке мембраны); а — доля открытых каналов; у — проводимость одиночного канала. По своей селективности электровозбудимые ионные каналы нервных и мышечных клеток подразделяются на натриевые, калиевые, кальциевые, хлорные. Селективность эта Не абсолютная: название канала указывает лишь ион, для которого данный канал наиболее проницаем. • Через открытые каналы ионы движутся по концентрационному и электрическому градиентам. Эти потоки ионов приводят к изменениям мембранного потенциала/что в свою очередь изменяет среднее число открытых каналов и соответственно величину ионных токов и т. д. Такая круговая связь важна для генерации потенциала действия, но. она делает невозможным количественную оценку зависимости ионных проводимостен от величины генерируемого потенциала. Для изучения этой зависимости применяется «метод фиксации потенциала». Сущность данного метода состоит в насильственном поддержании мембранного потенциала на любом заданном уровне. Так, подавав на мембрану ток, равный по величине, но обратный по знаку ионному току, проходящему через открытые каналы, и измеряя этот ток при различных Потенциалах, исследователи получают возможность проследить зависимость потенциала от ионных проводимостей мем- Внутренний потенциал 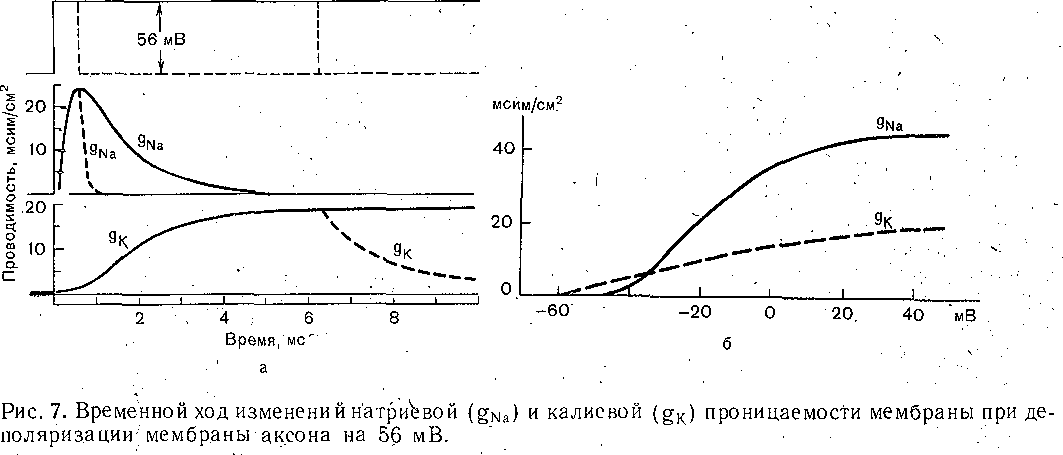 а,— сплошные^ инии показывают проницаемость при длительной деполяризации, а пунктирные — при ре поляризации мембраны через-0\В и 6,3 м'с;'б'>- зависимость пиковой величины натриевой (g^J и стационарного уровня калие.врй (gK) проницаемости о*т;мёмбраннВго потенциала. , 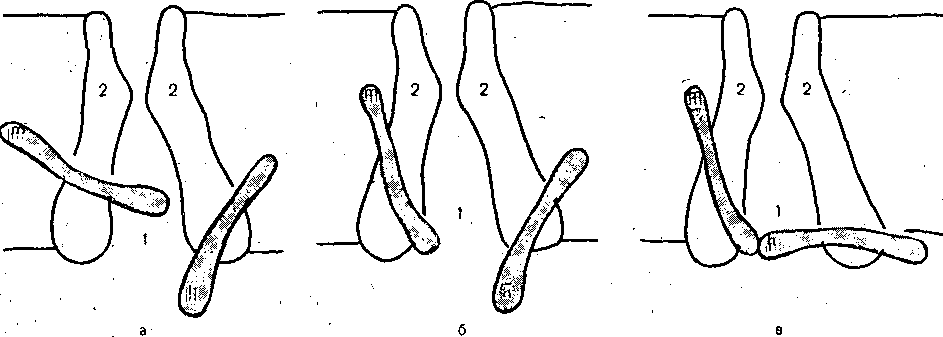 Рис. 8. Схематическое изображение эле ктро возбудимо го натриевого канала. Канал (1) образован макромолекулой белка 2), суженная часть которого соответствует «селективному фильтру». В канале имеются активационные (ш) и инактивационные (h) «ворота», которые управляются электрическим полем мембраны. При потенциале покоя (а) наиболее вероятным является положение «закрыто» Для активационных ворот и положение «открыто» для инактнвационных. Деполяризация мембраны (б) приводит к быстрому открыванию т-«ворот> и медленному закрыванию'11-«ворот», поэтому в начальный момент деполяризации обе пары «ворот» оказываются открытыми и через канал могут двигаться ионы в соответствии с их концентрационными и электрическими градиентами. При продолжающейся деполяризации (ii) ииактивационные «ворота» закрываются и капал переходит в состояние инактивации. браны. Чтобы из общего ионного тока, протекающего через мембрану, выделить его компоненты, соответствующие потокам ионов, например,,через натриевые каналы, используют химические агенты, специфически блокирующие всё другие каналы. Соответственным образом поступают при измерениях калиевого или кальциевого токов. На рис. 7 показаны изменения натриевой (gua) и калирвой (^к) проницаемости мембраны нервного волокна во время фиксированной деполяризации. Как. отмечалось, величины и gK отражают число одновременно открытых натриевых или калиевых каналов. Как видно, gNa быстро, за доли миллисекунды, достигла максимума, а затем медленно начала снижаться до исходного уровня. После окончания деполяризации способность натриевых каналов вновь открываться постепенно восстанавливается в течение десятков миллисекунд. 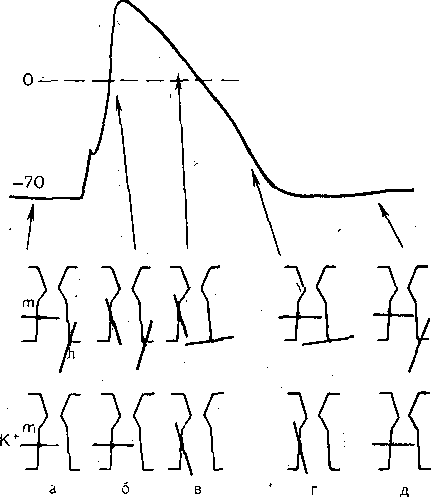 н50 мВ Потенциал действия Na*. Рис. 9. Состояние натриевых и калиевых каналов в различные фазы потенциалов действия (схема). Объяснение в тексте. Для объяснения такого поведения натриевых каналов высказано предположение о существовании в каждом канале двух типов «ворот» — быстрых активационных и медленных инактивационных. Как следует из названия, начальный подъем gNa связан с открыванием активационных ворот («процесс активации»), последующее падение во время продолжа- " ющейся деполяризации мембраны,— с закры- ' ванием инактивационных ворот («процесс инактивации»). На рис. 8, 9 схематически изображена организация натриевого канала, облегчающая понимание его функций. Канал имеет наружное и внутреннее расшит рення («устья») и короткий суженный участок, так называемый селективный фильтр, в котором происходит «отбор» катионов по их размеру и свойствам. Судя по размеру наибольшего проникающего через натриевый, канал катиона, отверстие фильтра не меньше 0,3—0,5 нм. При прохождении через фильтр ионы Ыа+ теряют часть своей гиДратной оболочки. Активационные (т) и инактивационные (/г) «воро та» расположены в области внутреннего конца натриевого канала, причем «ворота» /г'обращены в сторону цитоплазмы- К та кому заключению пришли на основании того факта, что приложение некоторых протеолитически* ферментов (проназы) к внутренней стороне мембраны приводит к ■устранению натриевой инактивации (разрушает /г-«ворота»), . ' В состоянии покоя «ворота» т закрыты, тогда как «ворота»h открыты. При деполяризации в начальный момент «ворота» тм h открыты — канал находится в проводящем состоянии. Затемгч инактивационные ворота закрываются — канал инактивируется. После окончания деполяризации «ворота» h медленно открываются, а «ворота» т быстро закрываются и канал возвращается в исходное покоящееся состояние. . , У Специфическим блокатором натриевых каналов является тетродотоксин,— соединение, синтезируемое в тканях некоторых видов рыб. и саламандр. Это соединение входит в наружное устье канала, связывается с какими-то'пока неидентифицирЬванными химическими группами и «закупоривает» канал. Используя радиоактивно меченный тетродотоксин, подсчитали плотность натриевых каналов в мембране. У различных клеток эта плотность варьирует от десятков до'десятков тысяч натриевых каналов на квадратный .микрон мембраны, ■ ' Функциональная организация калиевых каналов сходна с таковой натриевых каналов, различия лишь в их селективности и кинетике процессов активации и инактивации. Селективность калиевых каналов выше селективности натриевых: для Na+ калиевые каналы практически непроницаемы; диаметр их селективного фильтра около 0,3 нм. Активация калйевых каналов имеет примерно на порядок более медленную кинетику, чем активация натриевых каналов (см. рис. 7). На протяжении 10 мс деполяризации gK не обнаруживает тенденции к инактивации: калиевая "инактивация развивается только при многосекундной деполяризации мембраны. , Следует подчеркнуть, что-такие соотношения между процессами активации и инактивации калиевых каналов характерны только для нервных волокон. В мембране многих нервных и мышечных клеток существуют калиевые каналы, которые сравнительно быстро инактивИруются. Обнаружены также быстро активирующиеся калиевые каналы. Наконец, существуют калиевые каналы, которые активируются1 не мембранным потенциалом', а внутриклеточным Са2+, Калиевые каналы блокируются органическим катионом тетраэтиламмонием, а также аминопиридинами. ч Кальциевые каналы характеризуются медленной кинетикой процессов активации (миллисекунды) и инактивации (десятки и сотни миллисекунд). Их селективность определяется наличием в области наружного устья каких-то химических групп, обладающих повышенным сродством к двухвалентным катионам: Са2+ связывается с этими группами и только после этого проходит в полость канала. Для некоторых двухвалентных катионов сродство к указанным группам настолько велико, что, связываясь с ними, они блокируют движение Са + через канал. Так действует Кальциевые каналы мо гут быть блокированы также некоторыми органическими соединениями (верапамил, нифедипин), используемыми в.клинической практике для подавления повышенной электрической активности гладких мышц. ч Характерная особенность кальциевых каналов — их зависимость .от метаболизма и, в частности, от циклических нуклеотидов^(цАМФ и цГМФ), регулирующих процессы фосфорилирования и дефосфорйлирования белков кальциевых каналов. ' Скорость процессов .активации и инактивации всех ионных каналов увеличивается с возрастанием деполяризации мембраны; соответственно увеличивается До некоторой предельной величины ^число одновременно открытых каналов. МЕХАНИЗМЫ ИЗМЕНЕНИЯ ИОННОЙ ПРОВОДИМОСТИ ВО ВРЕМЯ ГЕНЕРАЦИИ ПОТЕНЦИАЛА ДЕЙСТВИЯ Известно, что восходящая фаза потенциала действия связана с повышением натриевой проницаемости. Процесс повышения развивается следующим образом. В отйет на начальную деполяризацию мембрдны, вызванную раздражителем, открывается лишь небольшое число натриевых каналов. Их открывание, однако, приводит к возникновению входящего внутрь клетки потока ионов Na+ (входящий натриевый ток), который увеличивает начальную деполяризацию. Это ведет к открыванию новых натриевых каналов, т. е. к дальнейшему повышению gNa соответственно входящего натриевого тока, а следовательно, к дальнейшей "деполяризации мембраны, что, в свою очередь, Обусловливает, еще большее повышение gNa и т: д. Такой круговой'лавинообразный процесс получил название регенеративной (т. е. самообновляющейся) деполяризации. Схематически он может быть изображен следующим образом: —>- Деполяризация мембраны Раздражитель Г 1 Входящий. Повышение натриевый -«—натриевой ток проницаемости Теоретически регенеративная деполяризация должна была бы завершаться повышением внутреннего потенциала клетки до величины равновесного нернстовского потенциала для ионов Ка": RT F р Na где Na^"—наружная, a Na^ — внутренняя: концентрация ионов Na+, 'При наблюдаемом соотношении10 ЈNa=+55 мВ. Эта величина является предельной для потенциала действия. В действительности, однако, пиковый потенциал никогда не достигает величины ЈNa,. во-первых, потому, что мембрана в момент пика потенциала действия проницаема не только для ионов Na+, >но и для ионов К+ (в значительно меньшей степени). Во-вторых,, подъему потенциала действия до величины Ема противодействуют восстановительные процессы, ведущие к восстановлению исходной поляризации (реполяризация мембраны). v Такими процессами являются снижение значенияgNll и повышение уровня Снижение^Na обусловлено тем, что активация натриевых каналов во время деполяризации сменяется их инактивацией; это приводит к быстрому уменьшению числа открытых натриевых каналов. Одновременно под влиянием деполяризации начинается медленная активация калиевых каналов, обусловливающая рост значения gK. Следствием увеличенияgK является усиление выходящего из клетки потока ионов К+ (выходящий калиевый ток). • . В условиях понижения связанного с инактивацией натриевых каналов, выходящий ток ионов К+ приводит к ре'поляризации. мембраны или даже к ее временной («следовой») гиперполяризации, как это имеет место, например, в гигантском аксоне кальмара (см, рис. 4). Реполяризация мембраны в свою очередь ведет к закрыванию калиевых каналов^ и, следовательно, ослаблению выходящего калиевого тока. Вйесте с тем под влиянием реполяризации происходит медленное устранение натриевой инактивации,: открываются инактивационные ворота и натриевые каналы возвращаются в состояние покоя. На рис. 9 схематически показано состояние натриевых и калиевых каналов в различные фазы развития потенциала действия. Все агенты, блокирующие натриевые каналы (тетродотоксин, местные анестетики и многие другие препараты), снижают, крутизну нарастания и амплитуду потенциала действия и тем в большей степени, чем выше концентрация этих веществ. • АКТИВАЦИЯ НАТРИЙ-КАЛИЕВОГО НАСОСА ' ПРИ ВОЗБУЖДЕНИИ Возникновение серии импульсов в нервном или мышечном волокне сопровождается обогащением протоплазмы Na+ и потерей К+. Для гигантского аксона кальмара диаметром 0,5 мм подсчитано, что во время одиночного нервного импульса через каждый квадратный микрон мембраны в протоплазму поступает около 20 ООО Na+ и столько же К+ покидает волокно, В итоге при каждом импульсе аксон теряет около одной миллионной общего содержания калия. Хотя эти потери.очень незначительны, при ритмическом следовании импульсов, суммируясь, они должны были бы привести к более или менее заметным изменениям концентрационных градиентов., Особенно быстро такие концентрационные сдвиги должны были бы развиваться в тонких нервных и мышечных волокнах и мелких-нервных клетках, обладающих малым по отношению к поверхности объемом цитоплазмы. Этому, однако,'-противодействует натриевый насос, активность которого возрастает при повышении внутриклеточной концентрации ионов Na+. Усиление работы насоса сопровождается значительным повышением интенсивности обменных процессов, поставляющих энергию для активного' переноса ионов Na+ и К+ через мембрану. Этолроявляется усилением процессов распада и синтеза АТФ и креатин- фосфата, увеличением потребления клеткой кислорода, повышением, теплопродукции и т. п. Благодаря работе насоса нарушенное при возбуждении неравенство концентраций Na+ и К+ по обе стороны мембраны полностью восстанавливается. Следует,, однако, подчеркнуть, что скорость выведения Na+ из цитоплазмы с помощью насоса относительно мала: она примерно в 200 раз ниже скорости движения этих ионов через мембрану ло концентрационному градиенту. 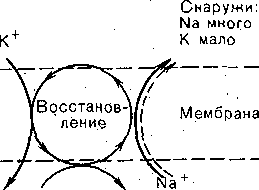 Обмен веществ Внутри: Na мало . К много Таким образом, в живой клетке существует'две системы движения ионов через мембрану (рис. 10). Один из них осуществляется по градиенту концентрации ионов и не требует затраты энергии, поэтому .его называют пассивным ионным транспортом. Он ответствен за возникновение потенциала покоя и потенциала действия и ведет в конечном итоге к выравниванию концентрации ионов по обе стороны клеточной мембраны: Второй тип движения ионов через мембрану, осуществляющийся против концентрационного градиента, состоит в «выкачивании» ионов натрия из цитоплазмы и -«нагнетании» ионов калия внутрь клетки. Этот тип ионного транспорта возможен лишь при условии затраты энергии обмена веществ. Его называют активным ионным транспортом. Он ответствен за поддержание постоянства разности концентраций ионов между цитоплазмой и омывающей клетку жидкостью. Активный транспорт — результат работы натриевого насоса, благодаря которому восстанавливается исходная разность ионных концентраций, нарушающаяся при каждой вспышке возбуждения. Рис. 10. Две системы транспорта ионов через мембрану. Справа — движение ионов Na+ и Кн" по ионным каналам во время возбуждения в соответствии с концентрационным и электрическим градиентами. Слева — активный транспорт ионов против концен- трационного градиента за счет энергии метаболизма («натриевый насос»). Активный транспорт обеспечивает поддержание и восстановление ионных градиентов, изменяющихся вр время импульсной активности. Пунктирной линией обозначена та часть оттока Na + , которая не исчезает при удалении из наружного раствора ионов К+ [Ходжкин А., 1965]. . .. МЕХАНИЗМ РАЗДРАЖЕНИЯ КЛЕТКИ (ВОЛОКНА) ЭЛЕКТРИЧЕСКИМ ТОКОМ В естественных условиях генерацию потенциала действия вызывают так называемы) местные токи, возникающие ме>кду возбужденным (деполяризованным) и покоящимс: участками клеточной мембраны. Поэтому электрический ток рассматривается как аде кватный раздражитель для возбудимых мембран и успешно используется в эксперимен тйх при изучении закономерностей возникновения потенциалов действия. Минимальную силу тока, необходимую и достаточную для инициации потенциал* действия, называют пороговой, соответственно раздражители большей и меньшей силь обозначают подпороговыми и сверхпороговыми. Пороговая сила тока (пороговый ток) в определенных пределах находится в обратной зависимости от длительности его дейст вия. Существует также некоторая минимальная крутизна нарастания силы Тока, ний<( которой последний утрачивает способность вызывать потенциал действия. Существуют два способа подведения'тока к тканям для измерения порога раздражения и следовательно, для'определения их возбудимости. При первом способе — внеклеточном — оба электрода располагают на поверхности'раздражаемой ткани. Условно принимают, что приложенный ток входит в ткань в области анода и выходит в области катода (рис. И). Недостаток этогс метода измерения порога заключается в значительном ветвлении тока: только часть его прохоркт через мембраны клеток, часть же ответвляется в. межклеточные щели. Вследствие этого при раздражении приходится применять ток значительно большей силы, чем необходимо для возникновения возбуждения.: ' - При втором способе подведения тока-к-клеткам — внутриклеточном —, микроэлектрод вводят в клетку, а обычный электрод прикладывают к поверхности ткани (рис> 12). В этом случае весь ток проходит через мембрану клетки, что позволяет точно определить наименьшую силу тока, необходимую для возникновения потенциала действия. При таком способе-раздражения отведение потенциалов производят с помощью второго внутриклеточного микроэлектрода. Пороговая сила тока, необходимая для возникновения возбуждения различных клеток при внутриклеточном раздражающем электроде,, равна 107 — Ю-9 А. . В лабораторных условиях и при проведении, некоторых клинических исследований для раздражения нервов и мышц применяют .электрические стимулы различной формы: прямоугольной, синусоидальйой, линейно и экспоненциально нарастающей, индукционные удары, конденсаторные разряды и т. п. , - Механизм раздражающего действия тока при,всех видах стимулов в принципе одинаков, однако в наиболее отчетливой форме он выявляется при использовании постоянного тока.. 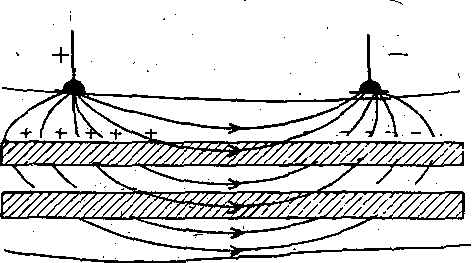 Рис. 11, Ветвление тока в ткани при раздражении через , наружные (внеклеточные) электроды (схема). : Осциллограф 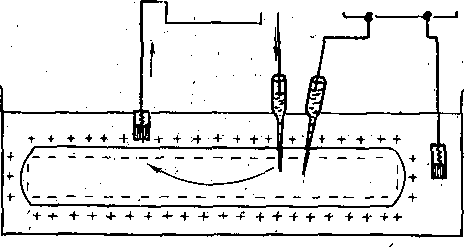 — ! Стимула-1 ' -f ГУсилитель л * тор Т7 пост, тона Рис. 12. Раздражение и отведение потенциалов через внутриклеточные микроэлектроды. Объяснение в тексте. 33 Мышечные волокна заштрихованы, между н^- ми — межклеточные щели. 2 Физиология человека ДЕЙСТВИЕ ПОСТОЯННОГО ТОКА НА ВОЗБУДИМЫЕ ТКАНИ Полярный закон раздражения При раздражении нерва иди мышцы постоянным током возбуждение возникает в юмент замыкания постоянного тока только под катодом, а в момент размыкания — олько под анодом. Эти факты объединяют под названием полярного закона раздраже- !ия, открытого Пфлюгеровд в 1859 г. Полярный закон доказывается следующими опы- ами. Умерщвляют участок нерва под одним из электродов, а второй электрод уста- [авливают на неповрежденном участке. Если с неповрежденным участком сопри- ;йсается. катод,-возбуждение .возникает в момент замыкания тока; если же катод гста на вливают на поврежденном участке, а анод — на неповреждённом, возбуждение юзникает только при размыкании' тока. Порог раздражения при размыкании, ;огда возбуждение возникает под анодом, значительно выцле, чем при замыкании, когда юзбуждение возникает под катодом. Изучение механизма полярного действия электрического тока стало возможным юлько после того, как был разработан описанный метод одновременного введения в тетки двух микроэлектродов: одного — для раздражения, другого— для отведения потенциалов. Было установлено, что потенциал действия возникает только в том случае, ^сли катод находится снаружи, а анод—• внутри клетки. При обратном расположении толюсов, т. е. наружном аноде и внутреннем катоДе, возбуждения при замыкании тока le возникает, как бы силен он ни был. 1 " ' г Прохождение через нервное или. мышечное волокно электрического тока прежде всего вызывает изменения мембранного потенциал^. В области приложения к поверхности ткани анода положительный потенциал на наружной стороне мембраны возрастает, т. е. происходит гиперполяризация, а в том случае, когда к поверхности приложен катод, положительный потенциал на наружной стороне мембраны снижается — возникает деполяризация. . ,. На рис. 13, а показано, что как при замыкании, так и при размыкании тока изменения мембранного потенциала нервного- волокна не возникают и не исчезают мгновенно, а плавно развиваются во времени. ' " Объясняется это тем,- что поверхностная мембрана живой клетки обладает свойствами конденсатора. Обкладками этого «тканевого конденсатора» служат наружная и внутренняя поверхности мембраны, а диэлектриком — слой липидов, обладающий значительным сопротивлением. Ввиду наличия в мембране каналов, через которые могут проходить ионы, сопротивление этого слоя не равно бесконечности, как в идеальном конденсаторе. Поэтомуу поверхностную мембрану клетки обычно уподобляют конденсатору с параллельно включенным сопротивлением, по которому может происходить утечка.зарядов (рис. 13, а). Временной ход изменений мембранного потенциала при включении и выключении тока (рис. 13,. б) зависит от емкости С и сопротивления мембраны R. Чем меньше произведение ДС— постоянная времени мембр'аны, тем быстрее при данной силе тока нарастает потенциал и, наоборот, большей величине RC соответствует меньшая скорость увеличения потенциала. Изменения мембранного потенциала возникают не только непосредственно в точках приложения к нервному волокну катода и анода постоянного тока^но и на некотором расстоянии от полюсов с той, однако, разницей, что их величина постепенно убывает по мере удаления от катода и анода. Объясняется это так называемыми кабельными свойствами нервного и мышечного волокон. Однородное нервное волокно в электрическом отношении представляет собой кабель, т. е. сердечник с низким удельным сопротивлением (аксоплазма)', покрытый изоляцией (мембраной), и помещенный в хорошо проводящую среду. Эквивалентная схема кабеля приведена на,рис. 13, б. При прог пускании, через некоторую точку волокна длительное время постоянного тока наблюдается стационарное состояние, при котором плотность тока и, следовательно, изменение мембранного' потенциала максимальны в месте приложения тока (т. е. непосредственно под катодом и анодом); с удалением от полюсов плотность тока и изменения потенциала на мембране экспоненциально уменьшаются по длине волокна. Поскольку рассматриваемые изменения мембранного потенциала в отличие от локального, ответа потенциала действия или следовых потенциалов не связаны с изменениями ионной проницаемости мембраны (т.-е. актйвным ответом волокна), их принято называть пассивными, 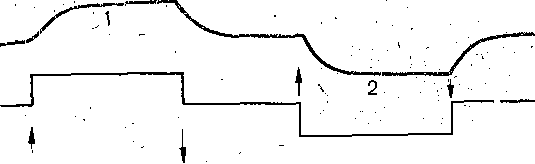 Потенциал Катод Анод Рис. 13. Простейшая электрическая схема, воспроизводящая электрические свойства мембраны (а и изменения мембранного потенциала под катодом и анодом постоянного тока.подпороговой си лы (б). а: С — емкость мембраны, R — сопротивление, Е — электродвижущая сила .мембраны в покое (потенциа; покоя).. Приведены средние значения R, С и Ё для мотонейрона, б — деполяризация мембраны (1) под катодом и гиперполяризация (2) под анодом при Ьрохождении через нервное волокно слабого подпороговогс тока. . ' или «электротоническимиизменениями мембранного потенциала. В чистом виде последние могу быть зарегистрированы в-условиях полной блокады ионных каналов химическими агентами. Разли чают кат- и анэлектротоническиё изменения потенциала, развивающиеся в области приложение соответственно катода и анода постоянного тока. - Критический уровень деполяризации - \ Регистрация изменений мембранного потенциала при внутриклеточном раздражении нервного или мышечного волокна показала, что потенциал действия возникает в. тот момент, когда деполяризация мембраны достигает критического уровня. Этот критический уровень деполяризации не зависит от характера примененного стимула, расстояния между электродами и т. п., а определяется исключительно свойствами самой мембраны. На рис. 14 схематически показаны изменения мембранного потенциала нервного волокна под влиянием длительного и-коротких стимулов различной силы. Во всех случаях потенциал действия возникает тогда, когда мембранный, потенциал достигает критической величины. Скорость, с которой происходит деполяризация мембраны, при прочих равных 4 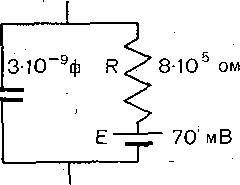 Наружная сторона Внутренняя сторона  1. Л_ о > условиях зависит лот силы раздражающего тока. При токе слабой силы деполяризация развивается медленно, поэтому . для возникновения потенциала действия стимул должен быть-большей длительности. В случае усиления раздражающего тока скорость развития деполяризации возрастает и. соответственно уменьшается минимальное время, необходимое для возникновения возбуждения. Чем быстрее .развивается деполяризация мембраны, тем меньше минимальное время, необходимое для генерации потенциала действиями наоборот. Локальный ответ , В механизме критической деполяризации мембраны наряду с пассивными существенную роль играют активные подпороговые изменения мембранного потенциала, проявляющиеся й форме так называемого локального ответа. Время Рис. 14. Изменение мембранного, потенциала до критического уровня деполяризации мембраны при действии раздражающего тока разной силы и длительности. Критический уровень показан пунктиром. Внизу — раздражающие стимулы, при воздействии которых получены, ответы А, Б и В. эис. 15. Локальный ответ нервного волокна. Б, В —изменения мембранного потенциала 1ервного волокна, вызываемые действием подпоро- ово'го тока короткой длительности/На кривых Б и 3 к пассивной деполяризаиии мембраны присоеди- шется и активная подпороговая деполяризация в |зорме локального ответа. Локальный ответ отде- 1ен от пассивных изменений потенциала пунктир- юй линией. При пороговой силе тока (Г) локальный ответ перерастает в потенциал действия "(его 5ершина на рисунке не показана). человека 1 УК1 5Л4 2 gr. ■ /V-«7, ' 34 ПРОВЕДЕНИЕ НЕРВНОГО ИМПУЛЬСА И НЕРВНО-МЫШЕЧНАЯ ПЕРЕДАЧА 88 ишшшш! 95 ВВЕДЕНИЕ 114 ОБЩАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ 117 частная физиология 148 центральной нервной Системы 148 НЕРВНАЯ РЕГУЛЯЦИЯ ВЕГЕТАТИВНЫХ ФУНКЦИЙ 210 гормональная регуляция физиологических функций 231 |