|
|
Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
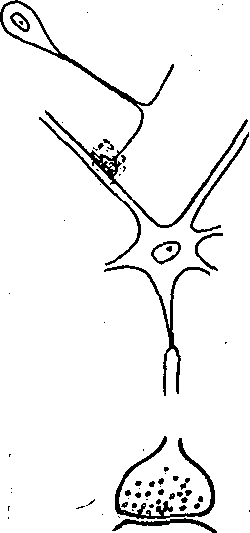
Структурные и функциональные особенности электрических, химических и смешанных синапсов. Для того чтобы потенциал действия, пр-иходящий в пресинаптическое окончание аксона, мог непосредственно возбудить постсинаптическую мембрану, т. е. вызвать в ней изменение мембранного потенциала, необходимо, чтобы значительная часть тока, текущего через пресинаптическую мембрану, могла входить в постсинаптическую клетку. Условием для такого вхождения тока является низкое сопротивление участка, связывающего обе клетки (они должны быть электрически связаны), и отсутствие шунтов, по которым пресинаптический ток мог бы ответвиться и не попасть на постсинаптическую мембрану.
Если синаптическая щель, разделяющая пре-"и постсинаптическую мембраны, широкая (как это имеет место в химических синапсах, где она составляет в среднем 10—-20 нм), подавляющая часть пресинаптического тока шунтируется'низким сопротивлением щели и лишь примерно 0,0001 часть его попадает на постсинаптическую мембрану. Эта величина слишком мала, чтобы вызвать ощутимый сдвиг мембранного потенциала постсинаптического нейрона. Поэтому в. синапсах с широкой синаптической щелью необходим другой механизм, способный изменить мембранный потенциал постсинаптической клетки. Таким- механизмом является выделение пресинапсом особых химических веществ — медиаторов, которые, воздействуя на специфические рецепторы Постсинаптической мембраны, способны изменять состояние ионных каналов постсинаптической мембраны. Изменение ионной проницаемости постсинаптической мембраны, в свою очередь, приводит к возникновению постсинаптического ионного тока, вызывающего падение напряжения на постсинаптической мембране — постсинаптический потенциал. Работа химического синапса схематически изображена на рис. 62, а. Таким образом, генератор постсинаптического тока находится непосредственно в постсинапти-
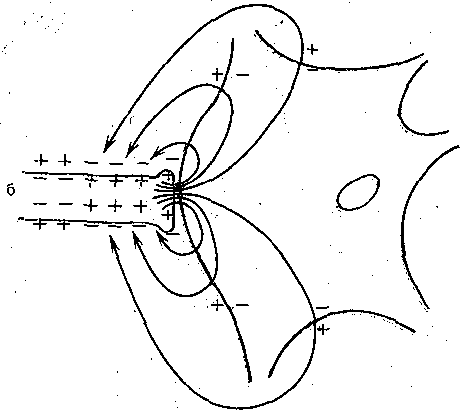
Рис. 62. Схема передачи возбуждения в химическом (а) и электрическом синапсе (б). Стрелками показано распространение электрического тока через мембрану пресинаптического окончания и постсинаптическую мембрану на нейрон.
ческой мембране и запускается химическим медиатором, выделяемым пресинаптическим окончанием. ;
В электрических синапсах ширина синаптической щели составляет всего 2—4. нм, что значительно меньше, чем в химических синапсах. Особенно важным является то, что в таких синапсах через синаптическую щель перекинуты мостики, образованные белковыми частицами. . Они представляют собой своеобразные каналы шириной 1—2 нм, пронизывающие пре- и постсинаптическую мембраны синапса. Благодаря существованию таких каналов, размеры которых позволяют переходить из клетки в клетку неорганическим ионам и даже небольшим молекулам, электрическое сопротивление в области такого синапса (получившего название щелевого или высокопроницаемого контакта) оказывается очень низким. Это позволяет пресинаптическому току распространяться на постсинаптическую клетку без угасания. Поэтому механизм работы электрического синапса сходен в общих чертах с механизмом распространения волны деполяризации по нервному или мышечному волокну, Электрический ток течет от возбужденной области к невозбужденной и там вытекает наружу, вызывая ее деполяризацию (рис. 62, б). В электрическом синапсе потенциал действия достигает пресинаптического окончания и далее течет через межклеточные каналы, вызывая деполяризацию постсинаптической мембраны, т. е. генерируя возбуждающий поетеинаптический потенциал (ВПСП). Важно подчеркнуть, что в электрическом синапсе генератор постсинаптического тока находится в пресинаптической, мембране, где возникает активный процесс — потенциал действия. Из нее он пассивно (электротонически) распространяется на мембрану постсинаптической клетки. Поэтому синапсы с электрическим механизмом передачи часто обозначают как электротонические. .
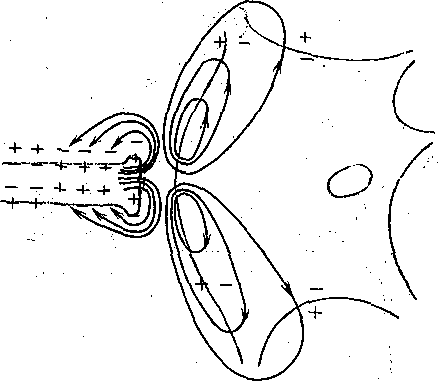
Структурная основа электрического синапса — высокопроницаемый щелевой контакт, обеспечивающий не только хорошую электрическую связь между нервными, клетками, но и взаимный обмен различными органическими молекулами диаметром 1—2 нм. Более крупные молекулы, например белки, ДНК и РНК через межклеточные каналы не .проходят. Однако и ограниченный обмен молекулами и ионами способен обеспечить определенную «метаболическую кооперацию» между нейронами, соединенными электрическими синапсами. - Хотя электрические синапсы немногочисленны в ЦНС высших животных, они широко, распространены в других возбудимых и невозбудимых тканях: в сердечной мышце, гладкой мускулатуре внутренних органов в печени, эпителиальной и железистых тканях.
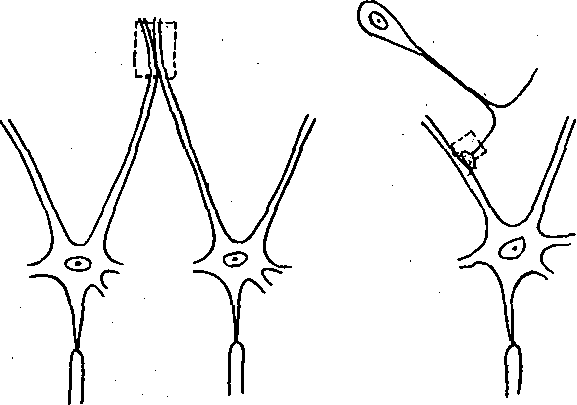
Рис. 63. Схема чисто электрического синапса между дендрита ми мото нейронов лягушки (а), смешанного синапса между центральными окончаниями первичного афферентного нейрона и мотонейроном лягушки (б) и химического синапса между центральными окончаниями первичного афферентного нейрона и мотонейрона кошки (в).
В некоторых межнейронных синапсах электрическая и химическая передача осуществляются параллельно благодаря тому, что щель между ripe- и постсинаптической мембранами имеет участки со структурой химического и электрического синапсов. Все 3 типа синапсов: электрический, химический и смешанный — схематически показаны на рис. 63. Обычно чисто электрические синапсы имеются между однотипными, близко расположенными нейронами, например между дендритами мотонейронов. Аксодендри- тические или аксосоматические синапсы, последовательно соединающие разные по функции и локализации нейроны, например первичные афферентные нейроны и мотонейроны, имеют химическую или смешанную природу.
Электрические и химические синапсы значительно отличаются друг от друга не только механизмом передачи, но и многими функциональными свойствами:
В синапсах с химическим механизмом передачи 11родолжй'тельность сипиитической задержки у теплокровных составляет 0,2—0,5 мс. В электрических синапсах синаптическая задержка, т. е. интервал между приходом импульса в пресинаптическое окончание и началом постсинаптического потенциала, отсутствует,
Химические синапсы отличаются односторонним проведением-, медиатор, обеспечивающий передачу сигналов, содержится только в пресинаптическОм звене. В электрических синапсах проведение чаще двустороннее, хотя геометрические особенности синапса делают проведение в одном направлении более эффективным. Кроме того, односторонность проведения в электрических синапсах может быть обеспечена полупроводниковыми свойствами мембраны.
Ввиду того что в химических синапсах возникновение постсинаптического потенциала обусловлено изменением ионной проницаемости постсинаптической мембраны, они эффективно обеспечивают как возбуждение, так и торможение постсинаптического нейрона. В электрических синапсах активный процесс развивается в пресинаптическом звене, и поскольку нервный импульс всегда представляет собой волну деполяризации,
электрические синапсы могут обеспечить передачу только одного процесса — возбуждения.
Химические синапсы значительно лучше, чем электрические, сохраняют следы предшествующей активности. Поэтому химическая передача значительно более подвержена модуляции под влиянием разных факторов.
Химические синапсы значительно более чувствительны к изменениям температуры, чем электрические, что имеет существенное значение для нервной системы пойкилотермных животных.-
Поскольку химический механизм синаптической передачи имеет значительно более широкое распространение, чем электрический, детальный анализ факторов, определяющих передачу сигналов в химических синапсах, особенно важен для понимания различных аспектов деятельности ЦНС в норме и патологии (а также действия на мозг различных фармакологических веществ и токсинов, пластических функций нервной системы и т. д.). Поэтому необходимо детально рассмотреть механизмы функционирования синапсов с химическим механизмом передачи, а именно высвобождение медиатора пресинаптическими окончаниями, химическую природу медиаторов, молекулярную и ионную структуру их действия на постсинаптическую мембрану нейронов, лежащую в основе синаптического возбуждения и торможения.
процесс высвобождения медиатора
Фактор, выполняющий медиаторнукх функцию, вырабатывается в теле нейрона и оттуда транспортируется в окончания его аксона, где в основном происходит его депонирование. Содержащийся в пресинаптических окончаниях медиатор должен выделиться в синаптическую щель, чтобы воздействовать на рецепторы постсинаптической мембраны, обеспечивая транссинаптическую передачу сигналов.
Еще до того, как были выяснены многие существенные особенности процесса высвобождения медиатора, было установлено, что пресинаптические окончания могут изменять состояние спонтанной секреторной активности. Выделяемые постоянно небольшие порции медиатора вызывают в постсинаптической клетке так называемые спонтанные миниатюрные постсинаптические потенциалы. Открытие спонтанного, т. е. не связанного с приходом нервного импульса, выделения медиатора помогло установить важнейшую особенность механизма его высвобождения— дискретный, квантовый характер. Дискретность процесса высвобождения выражается в том, что медиатор выходит из окончания не диффузно, не в виде отдельных молекул, а в форме многомолекулярных порций (или квантов), в каждой из которых содержится несколько тысяч молекул.
Постсинаптические эффекты, вызываемые в нервных клетках спонтанно выделяющимися квантами медиатора, наблюдаются особенно отчетливо в условиях, когда импульсная активность пресинаптических волокон искусственно подавлена, например, с помощью тетродотоксина — яда, избирательно блокирующего потенциалзависимые натриевые каналы мембраны, что устраняет возможность генерации потенциала действия. На рис. 64 показано, что после устранения импульсной активности спонтанные миниатюрные постсинаптические потенциалы продолжают возникать через нерегулярные интервалы времени. ■
Приходящий в пресинаптическое окончание нервный импульс резко увеличивает высвобождение квантов медиатора. Возникающий в результате одновременного высвобождения многих квантов постсинаптический ответ, таким образом, представляет собой вызванный постсинаптический потенциал. Деполяризация пресинаптической мембраны является необходимым условием для высвобождения медиатора. Установлено, что такая деполяризация будет неэффективной, если в окружающей нейроны среде отсутствуют ионы кальция.
Действительно, если изолировать участок мозга и перфузировать его искусственным раствором, то при сохранении обычного ионного состава перфузирующей жидкости синаптическая передача в изолированном мозге не будет отличаться от передачи в условиях целого мозга и нор-
^ujW
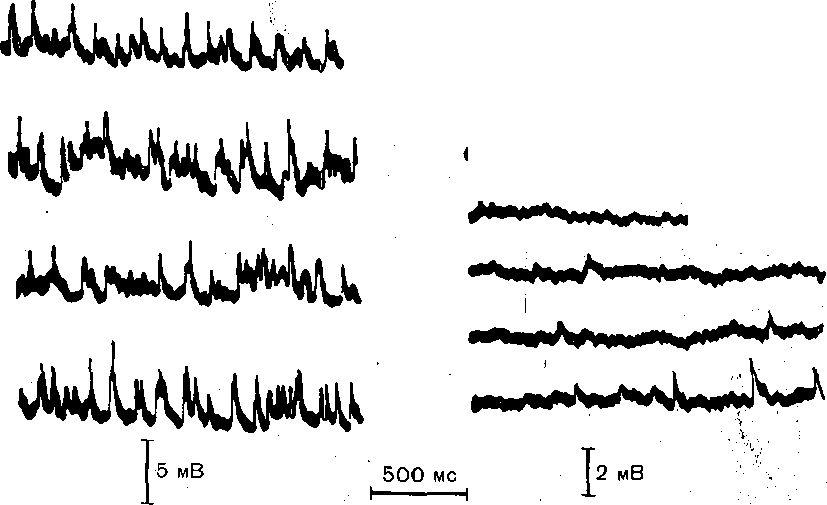
Рис. 64. Спонтанные миниатюрные постсинаптические потенциалы, зарегистрированные припомоцц внутриклеточного микроэлектрода в мотонейроне изолированного спинного мозга котенка.
а — при перфузии изотоническим раствором хлорида натрия; б — после выключения импульсной активное™ тетро до то кси но м.
мального кровообращения. Удаление из перфузата ионов Са2+ и особенно замена этих ионов на ионы Mg2+ или Мп2+, не влияя на спонтанное высвобождение квантов медиатора, прекращает высвобождение квантов медиатора нервными импульсами. Это особенно наглядно демонстрируют опыты на изолированном спинном мозге лягушки, поскольку здесь первичные афферентные волокнг образуют смешанные синапсы со спинальными мотонейронами и возбуждающие постсинаптические потенциалы, возникающие в мотонейроне при раздражении одного такого афферентного волокна, содержат электрический и химический компоненты (рис. 65). Электрический компонент, отражающий возникновение потенциала действия в пресинаптической окончании, не изменяется после удаления кальция из раствора. Напротив, медиаторный компонент полностью утрачивается. Таким образом, Са 2+ является необходимым для процесса высвобождения медиатора нервным импульсом. При отсутствии Са 2+ связь между деполяризацией пресинаптической мембраны и высвобождением медиатора (электросекреторная связь) нарушается. Роль кальция в этом .процессе связана с тем, что деполяризация, создаваемая нервными импульсами, приводит к активации потенциалзависимых кальциевых каналов пресинаптической мембраны. Ионы Са, поступая внутрь пресинаптического окончания, обеспечивают выход квантов Медиатора в синаптическую щель. Ионы Mg и Мп, блокируя потенциалзависимые кальциевые каналы мембраны, нарушают процесс высвобождения медиатора даже при наличии ионов Са в среде, окружающей клетки.
Ионы.Са также участвуют и в спонтанном выбросе квантов медиатора, так как факторы, способствующие увеличению концентрации Са 2+ внутри нервных окончаний, например некоторые метаболические ингибиторы, вызывают повышение частоты спонтанных миниатюрных потенциалов.
Дискретный, квантовый характер высвобождения медиатора нервным импульсом в синапсах ЦНС подтверждается результатами статистического анализа распределения амплитуд постсинапти- ческих потенциалов, вызываемых раздражением одиночного пресинаптического волокна.
97
В разных синапсах ЦНС эффекты, вызываемые в постсинаптической мембране одиночным квантом медиатора, и средний квантовый состав, т. е. число квантов медиатора, освобождаемых в среднем нервным импульсом, значительно варьируют. Так, в синапсах между окончаниями первичных афферентных волокон и мотонейронами спинного мозга величина деполяризации, вызываемая одним квантом медиатора, обычно составляет 50—100 мкВ, а число квантов, высвобождаемых окончаниями одного волокна на данном мотонейроне, обычно не превышает 5—10, а часто бывает значительно меньше. В синапсах между некоторыми клетками коры мозжечка средний квантовый состав может достигать нескольких сотен.
4 Физиология человека
Л Л
■ -0 гЭ]
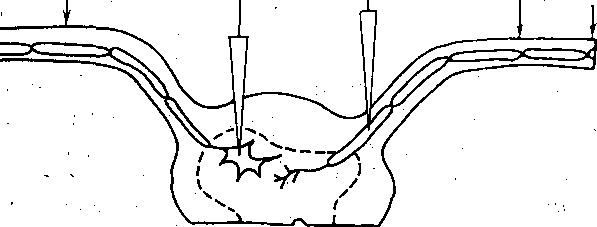
Рис.- 65. Схема опыта,, позволяющего регистрировать постсинаптические ответы в мотонейроне изолированного спинного мозга лягушки при раздражении одиночного пресинаптического волокна (а). Кривые (1, 2) иллюстрируют влияние.удаления кальция на химический компонент такого элементарного постсинаптического ответа.
а
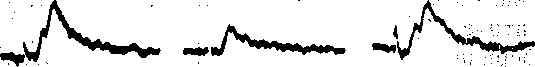 0 5 мВ
\
\
б
_J60 мВ 2 мс
Электросекреторная связь. Электронно-микроскопические исследования показали, то пресинаптические окончания всегда содержат синаптические пузырьки или везикулы, аждая из которых содержит один квант медиатора. Действительно, имеются;убедитель- ые биохимические данные, что вещества, рассматриваемые в качестве химических ме- иаторов, содержатся в синаптических пузырьках. Более того, расчеты количества гедиатора, содержащегося в одном пузырьке, и количество молекул медиатора, необхо- ямых для создания постсинаптического эффекта, аналогичного действию одного кванта, овпадают. Таким образом, совокупность имеющихся данных свидетельствует о том, что; ак спонтанные миниатюрные постсинаптические потенциалы, так и постсинаптические отенциалы обусловлены выходом в синаптическую щель медиатора, содержащегося синаптических пузырьках. Этот процесс (экзоцитоз) заключается в том, что пузырек, одойдя к внутренней поверхности мембраны пресинаптического окончания при наличии la 2+, сливается с пресинаптической мембраной. В результате происходит опорожнение узырька в синаптическую щель. После спадения пузырька окружающая его мембрана ключается в мембрану пресинаптического окончания, увеличивая его поверхность; \ дальнейшем (в результате Процесса эндоцитоза) небольшие участки пресинаптической 1ембраны впячиваются внутрь, вновь образуя пузырьки, которые впоследствии, снова пособны включать медиатор и вступать в цикл его высвобождения. Участие Са 2+ в процессе высвобождения медиатора нервным импульсом определяет 1ЯД важных специфических особенностей работы синапсов с химическим механизмом [ередачи. Такое характерное свойство химических синапсов, как синаптическая задерж- :а, определяется главным образом временем, необходимым для вхождения Са 2+ внутрь [ресинаптического окончания. Накопление Са 2" внутри пресинаптического окончания s результате предшествующего поступления улучшает эффективность работы химическо- •о синапса. Если интервал между последовательным возникновением потенциалов 1ействия в пресинапсе невелик, каждый последующий потенциал вызывает высвобожде- ше большего числа квантов медиатора, что проявляется увеличением амплитуды юстсинаптических потенциалов. Это явление временного облегчения или потенциации ложно связать с накоплением Са 2+ в пресинаптическом окончании. Такую же природу шеет и посттетаническая или постактивационная потенциация: увеличение числа кван- х>в медиатора, высвобождаемых нервным импульсом, после предшествующего ритми ческого раздражения. Посттетаническая потенциация может длиться от нескольки: минут до многих часов (в синапсах гиппокампа) и играть важную роль в пластически: изменениях функции синапсов. ХИМИЧЕСКИЕ МЕДИАТОРЫВ ЦНС медиаторную функцию выполняет не одно, а большая групп^ разнородны: химических веществ. Список вновь открываемых химических медиаторов неуклонш пополняется. Чаще всего химическими медиаторами являются вещества с небольшой от.носитель ной молекулярной массой. Однако и высокомолекулярные соединения, такие, как поли пептиды, также способны выполнять роль химических передатчиков в ряде центральны: и периферических синапсов. Основным критерием медиаторной функции веществ является его наличие в соот ветствующих пресинаптических окончаниях, способность высвобождаться под влияние? нервного импульса, а также идентичность молекулярных и ионных механизмов действи) на постсинаптическую мембрану вещества, высвобождаемого нервным импульсом и при кладываемого искусственно к пост си н а пти чес кой мембране. В противоположность многим периферическим структурам, где процесс идентифика ции медиатора по указанным выше критериям может быть произведен сравнительн< просто, ЦНС построена из негомогенных диффузно расположенных популяций нервны: клеток и окончаний. Это вносит значительные трудности в обнаружение выделяемой медиатора, который, прежде чем появиться на поверхности мозга или в спинномозгово1 жидкости, должен диффундировать,на большие расстояния. Помимо этого, в централь ных структурах трудно добиться избирательной стимуляции определенной гомогенно! группы нейронов или волокон, так же как трудно подводить предполагаемый медиато] к определенным нервным клеткам, не оказывая влияния на соседние нейроны. Именн< поэтому;природа химических медиаторов, во, многих синапсах ЦНС до сих пор оконча тельно не установлена. Тем не менее выявлен ряд веществ, играющих роль медиаторо! синаптического возбуждения и торможения в ЦНС млекопитающих и человека. К ним относятся: ацетилхолин; катехоламины: адреналин, норадреналин, дофамин серотонин: 5-гидрокситриптамин; нейтральные аминокислоты, глутаминовая, аспара гиновая кислоты; кислые аминокислоты: глицин, гамма-аминомасляная кислот* (ГАМК); полипептиды: вещество Р, энкефалин, соматостатин и др.; другие вещества АТФ, гистамин, простагландины. Согласно принципу Дейла, каждый нейрон во всех своих синаптических окончания: выделяет один и тот же медиатор. Поэтому принято, обозначать нейроны по.типу медиато ра, который выделяют их окончания. Нейроны, освобождающие ацетилхолин, называю' холинергическими, серотонин — серотонинергическими и т. д. Этот же принцип може' быть использован для обозначения, различных химических синапсов. Иными словами различают холинергические, серотонинергические и другие синапсы. |
|
|
 Скачать 7.39 Mb.
Скачать 7.39 Mb.