Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
 Скачать 7.39 Mb. Скачать 7.39 Mb.
|

|
Глава 7 НЕРВНАЯ РЕГУЛЯЦИЯ ВЕГЕТАТИВНЫХ ФУНКЦИЙ Со времени французского физиолога Бита — с начала XIX столетия — функции организма разделяют на ашшальШе, или соматические, и вегетативные. К анимальным, или соматическим, функциям относится восприятие внешних раздражений и двигательные реакции, осуществляемые скелетной мускулатурой. Вегетативными функциями называют те, от которых зависит осуществление обмена веществ в целостном организме (пищеварение, кровообращение, дыхание, выделение и т. д.), а'Таюке рост и размножение. В соответствии с этим разделением функций различают соматическую и вегетативную нервную систему. Соматическая нервная система обеспечивает ёкстероцептивные сенсорные и моторные функции организма. Вегетативная нервная система обеспечивает регуляцию деятельности внутренних Органов, сосудов и потовых желез, а также трофическую иннервацию скелетной •мускулатуры, рецепторов и самой нервной системы. Вегетативная нервная система отличается от соматической локализацией своих ядер в ЦНС, очаговым выходом волокон из'мозга, отсутствием сегментарности их распределения на периферии и малым диаметром волокон. Помимо этого, для вегетативной нервной системы характерно, что ее волокна, направляющиеся из мозга к внутренним органам, обязательно прерываются в периферических вегетативных ганглиях, образуя синапсы на нейроНах, расположенных в этих ганглиях. Аксоны ганглионарных нейронов оказывают влияние на внутренние органы. Ганглии являются не только образованиями, передающими импульсы,из ЦНС на органы и ткани. Они представляют собой вынесенные на периферию рефлекторные центры, способные регулировать функции внутренних органов в известной мере независимо; от ЦНС посредством периферических рефлексов, замыкающихся в этих ганглиях. Вегетативная нервная система делится на симпатический и парасимпатический отделы. Они отличаются по локализации центров в мозге, по характеру влияний на внутренние органы, а также тем, что ганглии парасимпатического отдела расположены в самих иннервируемых ими органах (интрамурально), в то время как ганглии симпатического отдела локализованы в пограничном стволе (truncus sympaticus). Выделяя регуляцию вегетативных функций, надо отметить, что в целостных реакциях организма сенсорные, моторные и вегетативные компоненты тесно связаны между собой. • . Соматические компоненты реакций организма, осуществляемые скелетной мускулатурой, з отличие от вегетативных могут быть произвольно вызваны, усилены или заторможены; они находятся в течение всего хода реакции под контролем сознания, Вегетативные же компоненты, как'правило, произвольно не контролируются. На этом основании вегетативную нервную систему называют автономной, или непроизвольной.. Однако представление об. автономности вегетативной нервной системы является весьма условным. Многочисленные опыты К. М. Быкова и сотрудников, показавшие возможность ус- ловнорефлекторной регуляции всех внутренних органов и всех вегетативных функций, позволяют считать, что кора больших полушарий регулирует деятельность всех органов, иннервированных вегетативной нервной системой, и координирует их деятельность в соответствии с текущими потребностями организма, в зависимости от характера его реакций при,изменениях внешней и внутренней среды организма. Это обстоятельство свидетельствует о принципиальной возможности произвольного управления вегетативными функциями, что удается осуществить после специальной целенаправленной тренировки (например, по системе индийских йогов). ОБЩИЙ ПЛАН СТРОЕНИЯ И ОСНОВНЫЕ ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ Центры вегетативной нервной системы расположены в мозговом стволе и спинном мозге. 1. В среднем мозге находятся мезэнцефальные центры парасимпатического отдела вегетативной нервной системы; вегетативные волокна от них идут в составе глазодвигательного нерва. 2. В продолговатом мозге расположены бульварные центры парасимпатического отдела нервной системы; эфферентные волокна от них проходят в составе лицево-4 го, языкоглоточного и блуждающего нервов. 3. В грудных и поясничных сегментах спинного мозга (от I грудного до II — IV поясничного) находятся тораколюмбальные центры симпатического отдела вегетативной нервной системы: вегетативные волокна от них выходят через передние корешки спинномозговых сегментов вместе с отростками моторных нейронов. 4, В крестцовых сегментах спинного мозга находятся сакральные центры парасимпатического отдела, вегетативной нервной системы, волокна от них идут в составе тазовых нервов. - Таким образом, центры вегетативной Нервной системы расположены в четырех отделах ЦНС. Ядра, находящиеся в мезэнцефальном, бульбарном и сакральном отделах,.образуют,парасимпатическую часть вегетативной нервной системы, а находящиеся в тора- колюмбальном отделе — ее симпатическую часть. Все уровни вегетативной нервной системы подчинены высшим, вегетативным центрам, расположенным в промежуточном мозге—в гипоталамусе и полосатом теле. Эти центры координируют функции многих органов и систем организма. Они в свою очередь подчинены коре больших полушарий, которая обеспечивает целостное, реагирование, организма, объединяя его соматические и вегетативные функции в единые акты/поведения. Симпатические нервные волокна имеют значительно более широкое распространение, чем парасимпатические. Симпатические нервы иннервируют фактически все органы, и ткани организма; напротив, парасимпатические же нервы не иннервируют скелетную мускулатуру, ЦНС, большую часть кровеносных сосудов и матку. . Распространение волокон парасимпатического отдела вегетативной нервной системы представлено на рис. 104. Ко многим органам парасимпатические волокна проходят в составе блуждающих нервов, которые иннервируют бронхи, сердце, пищевод, желудок, печень, тонкий.кишечник, поджелудочную железу, надпочечники, почки, селезенку, часть толстого отдела кишечника. 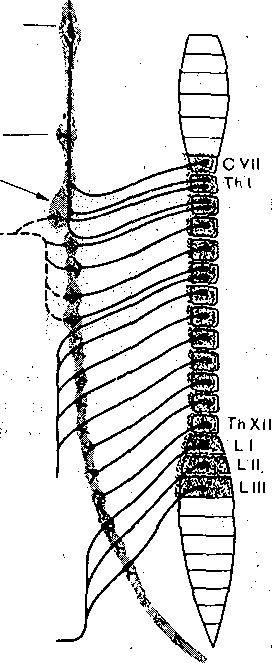 Верхней (лейный ганглий Средний шейный ганглий Звездчатый ганглий Симпатичесний ствол. 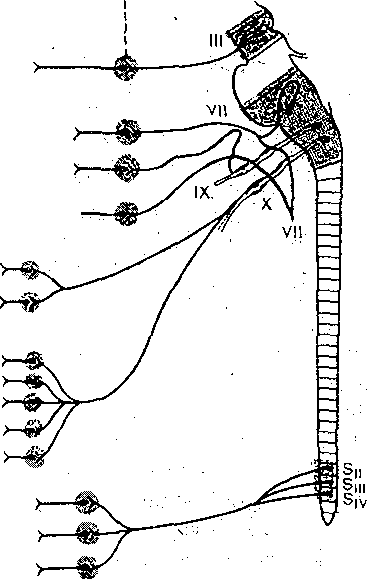 Ресничный ганглий ' < Слезная железа Околоушная железа Подчелюстная железа > Желудок Тонкие ницжи Печень Поджелудочная железа Почки Толстые мишки Мочевой пузырь Половые органы Зрачок Ветви к сердцу I и бронхам | ., Сердце Бронхи ролнечное сплет.екие Верхний' бры— точный узел Нижний брыжеечный узел ' I Рис. 104. Парасимпатический отдел вегетативной нервной Выделены: средний мозг, от Которого берут начало парасимпатические, ^'одцщ^^оходяшиё"в составе."ivia- зодвирательного нерва. (III); продолговатый мозг, от которого отходят па(раещпатичес.кие волокна в. составе лицевого {VII), языкоглоточного ОХ) и блуждающего (X) нервор1; крестцовыйбҐдел спинного мозга, от которого берет начало тазовый нерв. ' : \ Рис. 105. Симпатический отдел вегетативной нервной системы (схема). . Сплошные Линии —- преганглиоизрные волокна,- пунктирные — постганглиоиарные. Выделена тораколюм- бальная часть спинного мозга (от УП шейного до Ш поясничного сегмента); откуда берут начало прегангли- онарные симпатические волокна. \ .'' Распространение волокон симпатического отдела вегетативной нервной системы показано на рис. 105. Верхние сегменты симпатического отдела вегетативной нервной системы посылают свои волокна через верхний шейный симпатический узел к органам головы; следующие сегменты посылают их через нижележащие симпатические узлы к органам грудной полости и верхним конечностям; далее следует ряд грудных сегментов, посылающих врлокна через солнечное сплетение и верхний брыжеечный узел к органам брюшной полости, и, наконец, от поясничных сегментов волокна 'направляются через нижний брыжеечный узел в основном к органам малого таза и нижним конечностям. ДВУХНЕЙРОННАЯ СТРУКТУРА ЭФФЕРЕНТНЫХ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ПУТЕЙ Периферическая часть всех эфферентных симпатических и парасимпатических нервных путей построена из двух последовательно расположенных нейронов. Тело первого нейрона находится в ЦНС, его аксон Направляется на периферию и оканчивается в том или ином нервном узле. Здесь находится тело второго нейрона, на котором аксон первого нейрона образует синаптические окончания. Аксон второго нейрона иннервирует соот ветствующий орган. Волокна первого нейрона называют преганглионарными, второго — постганглионарными. Двухнейронная структура периферических эфферентных симпатических и парасимпатических путей является типичным признаком, отличающим их от соматических нервных волокон. На пути вегетативных нервов после выхода их из ЦНС, как правило, имеется только один перерыв нервного волокна, т. е. один синапс. Из этого правила, однако, имеются некоторые исключения. Так, постганглионарные симпатические волокна, идущие к гладким мышцам желудочно-кишечного тракта, преимущественно оканчиваются не на мышечных волокнах, а на парасимпатических ганглиозных клетках, находящихся в стенке желудка и кишок. По-видимому, они снижают активность этих клеток и таким путем оказывают тормозящее влияние на гладкую мускулатуру. В данном случае, следовательно, имеется трех- нейронная структура периферического пути. Исключением из отмеченного выше правила является также тот факт, что хромаффинные клетки мозгового слоя надпочечников иннервированы не постганглионарными, а преганглионарными симпатическими волокнами. Хромаффинные клетки, образующие под влиянием импульсов, поступающих к ним по симпатическим волокнам, адреналин, как бы заменяют постганглионарный нейрон, с которым они имеют общее происхождение. В данном случае имеется однонейронная структура эфферентного симпатического пути. ГАНГЛИИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ Ганглии симпатического отдела вегетативной нервной системы в зависимости от их локализации разделяют на вертебральные (иначе их называют паравертебральными) и превертебральные. Вертебральные симпатические ганглии расположены по обе стороны позвоночника, образуя два пограничных ствола (их называют также симпатическими цепочками). Вертебральные ганглии связаны со спинным мозгом нервными волокнами, которые образуют белые соединительные ветви — rami communicantes aibi. По ним к ганглиям идут преганглионарные волокна от нейронов, тела которых расположены в боковых рогах тораколюмбального отдела спинного мозга (рис. 106). Аксоны постгангли- онарных симпатических нейронов направляются от узлов к периферическим органам либо по самостоятельным нервным путям, либо в составе соматических нервов. В последнем случае они идут от узлов пограничных стволов к соматическим нервам в виде тонких серых соединительных веточек — rami communicantes grisei (серый их цвет обусловлен тем, что постганглионарные симпатические волокна не имеют миелиновых оболочек). В ганглиях пограничного ствола прерывается большинство симпатических преган- глионарных нервных волокон; меньшая их часть проходит через, пограничный ствол без перерыва и прерывается в превертебральных ганглиях. Превертебральные ганглии распространяются на большем, чем ганглии пограничного ствола, расстоянии от позвоночника; вместе с тем они находятся в некотором отдалении и от иннервируемых ими органов. К числу превертебральных ганглиев относят солнечное сплетение, верхний и нижний брыжеечные узлы. В них прерываются симпатические преганглионарные волокна, прошедшие без, перерыва узлы пограничного ствола. Ганглии парасимпатического отдела вегетативной нервной системы расположены внутри органов или вблизи них (ресничный узел gangl. ciliare, ушной узел — gangl. oticuni'H некоторые другие). Аксон первого парасимпатического нейрона, находящегося в среднем мозге, продолговатом мозге или в сакральном отделе спинного мозга, доходит до иннервируемого органа не прерываясь. Второй парасимпатический нейрон расположен внутри этого органа или в непосредственной близости от него — в прилежащем узле. Внутриорганные волокна и ганглии образуют сплетения, богатые нервными клетками, расположенные в мышечных стенках многих внутренних органов, например сердца, бронхов, средней и нижней третей пищевода, желудка, кишечника, желчного пузыря, мочевого пузыря, а также в железах внешней и внутренн(ей секреции. 161 Вегетативные ганглии играют важную роль в распределении и распространении проходящих через них нервных влияний. Число нервных клеток в ганглиях в несколько раз (в верхнем шейном симпатическом узле — в 32 раза, в ресничном узле — в 2 раза) 6 Физиология человека больше числа приходящих к ганглию пре- ганглионарных волокон. Каждое из этих волокон сильно ветвится и образует синапсы на многих клетках ганглия. Поэтому нервные импульсы, поступающие по преганглионарному волокну в. ганглий, могут оказывать влияние на' большое число ганглионарных нейронов и, следовательно, на еще большее число мышечных и железистых клеток иннервируемого органа. Таким образом достигается расширение зоны влияния преганглионарных волокон. На каждом ганглионарном нейроне имеются синапсы, образованные многими преганглионарными волокнами. С этой конвергенцией связано явление пространственной суммации нервных импульсов. Если раздражать одно преганглио- нарное волокно стимулами подпороговой силы, то в постганглионарных волокнах не возникает потенциалов действия. Если же раздражать несколько преганглионарных волокон стимулами той же силы, какая применялась для раздражения одного волокна, то обнаруживается потенциал действия в постганглионарных волокнах в- результате пространственной суммации постсинаптических возбуждающих потенциалов. При раздражении преганглионарных нервных волокон отчетливо обнаруживаются также явления временной суммации нервных импульсов. Раздражение одиночным стимулом, как правило, не дает эффекта даже при большой силе раздражения. Раздражение ритмическими стимулами преганглионарных волокон вызывает возбуждение постганглионарных нейронов вследствие временной суммации постсинаптических возбуждающих потенциалов. Одностороннее проведение нервных импульсов в межнейронных синапсах, перекрытие зон влияния отдельных входящих в узел преганглионарных волокон, наличие временной и пространственной суммации й окклюзии показывают, что нейроны и синапсы ганглиев вегетативной нервной системы обладают такими же свойствами, что нейроны и синапсы ЦНС. 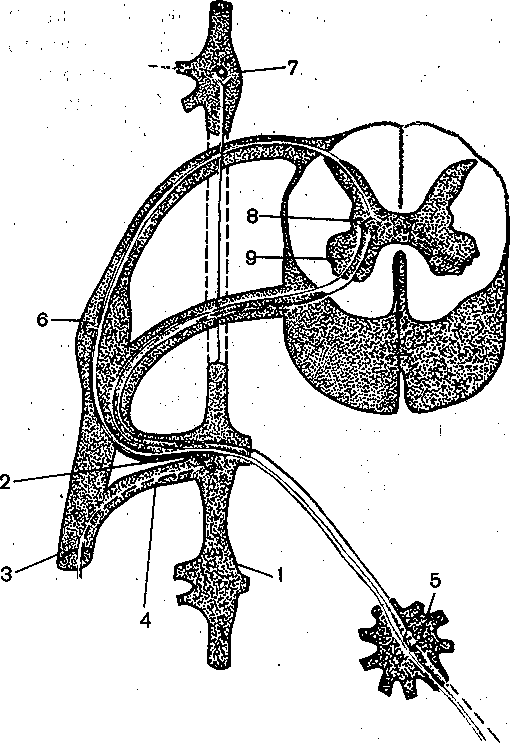 Рис. 106. Связь симпатических ганглиев со спинным мозгом. .
пульсов, проходящих по сосудосуживающим нервным волокнам, не бывает чаще 6- 8 имп/с. Частый ритм возбуждений преганглионарных волокон, превышающих частот; естественных импульсов, возникающих в нейронах вегетативной нервной системы, ча стично блокируется в синапсах ганглиев и постганглионарные волокна возбуждаются i более редком ритме. Таким образом, обнаруживается трансформация ритма нервны; импульсов. Частота стимуляции свыше 100 в секунду вызывает полную блокаду прове дения через синапс. Ганглии вегетативной нервной системы являются вынесенными на периферию ре флекторными центрами. ■ Поводом в пользу признания вегетативных ганглиев рефлекторными центрами явля ются морфологические наблюдения А. С. Догеля, который еще в конце прошлого столе тия описал в нервных сплетениях, находящихся в желудке и кишечнике, три типа нервны: клеток, отличающихся по своей форме. Одни, из этих клеток он считал рецепторными другие ■— моторными, третьи — вставочными. Между этими типами нервных клеток име ются синапсы, так что возможна передача влияний от рецепторного нейрона к эффектор ному. Вескими аргументами в пользу наличия рецепторных нейронов в периферически; нервных ганглиях являются обнаруженные факты сохранности афферентных, вста вочных и эфферентных нейронов и идущих от них нервных волокон в пересаженное собаке сердце другой:собаки. Если бы Эти рецепторы,чнервные клетки и нервные во локна относились к нейрону, тело которого находится в ЦНС, точнее за пределамг пересаженного сердца, то должно было произойти их перерождение. Однако рецепто- ры, нервные клетки, нервные волокна, синаптические.контакты в пересаженном сердце сохраняются. Они образуют внутрисердечную нервную систему, организованнук по рефлекторному принципу. Эта система может регулировать работу сердца путем ин- тракардиальных периферических рефлексов. В последние годы получены экспериментальные данные-о большом количестве «местных» периферических рефлексов, осуществляемых интрамуральными вегетативными ганглиями. Посредством таких рефлексоЕ регулируется деятельность сердца, перистальтика кишечника, осуществляется взаимосвязь разных отделов желудка и некоторых других органов. Периферические рефлексь: осуществляются превертебральными ганглиями (И. А, Булыгин). Это доказано при отведении потенциалов действия от одних нервов, отходящих от солнечного сплетения, и.раздражении других нервов. Однако рефлекторная функция, вероятно, осуществляется не всеми вегетативными ганглиями. Так, до сих пор не обнаружена такая деятельность в шейных симпатических ганглиях. Эфферентные нейроны вегетативных ганглиев получают импульсы из ЦНС (по преганглионарным вегетативным волокнам) и от внутриорганных рецепторов, образованных дендритами афферентных нейронов. Таким образом, эфферентные интрамураль- ные нейроны представляют собой общий конечный путь для импульсов внутриорганного и экстраорганного (центрального) происхождения. Наличие «местных» механизмов нервной регуляции функций внутренних органов, которая осуществляется с помощью периферических рефлек'сов ганглиями вегетативной нервной системы, внутриорганными и внеорганными, имеет большое физиологическое значение. В результате ЦНС освобождается от переработки избыточной информации. Кроме того, периферические рефлексы увеличивают надежность регуляции физиологических функций. Эта регуляция может осуществляться и после выключения связи органов с ЦНС-, 1 Вегетативные центры ЦНС получают информацию о состоянии внутренних органов от интерорецепторов по дендритам биполярных афферентных нейронов, расположенных в межпозвоночных узлах, и по ветвям аксонов интрамуральных афферентных нейронов. Импульсы, поступающие по этим путям в ЦНС, вызывают рефлекторные ответы не только вегетативной, но и соматической нервной системы. Они могут включать также сложные поведенческие реакции организма. Из сказанного ясно, что внутренние органы обладают богатой чувствительной иннервацией, обеспечивающей деятельность как периферических вегетативных рефлексов, так и реакций, осуществляемых вегетативными центрами мозга. Поступающая в ЦНС информация о состоянии внутренних органов необходима для возникновения мотиваций и, следовательно, участвует-в формировании сложных поведенческих реакций организма. Причиной таких реакций являются изменения не окружающей, а внутренней среды. Эти реакции направлены на удовлетворение той или иной биологической потребности организма. ТОНУС ВЕГЕТАТИВНЫХ ЦЕНТРОВ Многие центры вегетативной нервной системы постоянно находятся в состоянии активности, вследствие чего иннервированные ими органы получают от них возбуждающие или тормозящие импульсы непрерывно. Так, например, перерезка на шее собаки обоих блуждающих нервов влечет за собой учащение сердечных сокращений, так как при этом выпадает тормозящее влияние, постоянно оказываемое на сердце ядрами блуждающих нервов, находящимися в состоянии тонической активности. Односторонняя перерезка на шее кролика симпатического нерва вызывает расширение сосудов уха на стороне перерезанного нерва, так как сосуды лишаются вазоконстрикторного тонического влияния. При раздражении периферического отрезка перерезанного нерва в ритме 1—-2 имп/с восстанавливается тот ритм сердечных сокращений, который имел место до перерезки блуждающих нервов, или та степень сужения сосудов уха, которая была при целости симпатического нерва. Тонус вегетативных центров обеспечивается и поддерживается афферентными нервными сигналами, приходящими от рецепторов внутренних органов и отчасти от экстеро- рецепторов, а также в результате воздействия на центры разнообразных факторов крови и спинномозговой жидкости. СВОЙСТВА ВОЛОКОН ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ Преганглионарные волокна вегетативной нервной системы принадлежат к типу В; они имеют диаметр 2-—3,5 мкм (реже 5 мкм) и обладают тонкой миелиновой оболочкой. Постганглионарные волокна относятся к типу С; они имеют диаметр не более 2 мкм. Большая их часть не имеет миелиновой оболочки. ' Вегетативные, особенно постганглионарные, волокна отличаются малой возбудимостью: для их раздражения требуется большее напряжение электрического тока, чем для раздражения моторных волокон, иннервирующих скелетные мышцы. Скорость распространения по ним нервных импульсов мала: у млекопитающих она составляет в преганглионарных волокнах от 3 до 18 м/с, а в постганглионарных — от Г до 3 м/с. Чем тоньше волокно, тем больше его реобаза и хронаксия (т. е. меньше возбудимость), продолжительнее рефрактерность, меньше лабильность и медленнее скорость проведения импульсов. Потенциалы действия в симпатических и парасимпатических нервных волокнах отличаются большей длительностью, чем потенциалы действия соматических нервных волокон. Они сопровождаются в преганглионарных волокнах длительным следовым положительным потенциалом, а в постганглионарных волокнах —- следовым отрицательным потенциалом, переходящим в продолжительную (до 300 мс и более) следовую гиперполяризацию. ПЕРЕДАЧА ИМПУЛЬСОВ В СИНАПСАХ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ Медиатором, образующимся в окончаниях парасимпатических нервов, а также симпатических вазодилататоров и симпатических нервов потовых желез, является ацетилхолин; медиатором, образующимся в окончаниях постганглионарных симпатических нервов (за исключением нервов потовых желез и симпатических вазодилататоров),— норадреналин (адреналин, лишенный одной метильной группы). •Медиаторы, образующиеся в окончаниях вегетативных нервных волокон, действуют на йннервируемые ими клетки дольше по сравнению со временем действия медиатора (ацетилхолина) в окончаниях соматических нервов. По-видимому, это объясняется меньшей' активностью ферментов, разрушающих медиатор. Медиаторы образуются также тёрминалями преганглионарных волокон в синапсах ганглиев вегетативной нервной системы. Первые доказательства этого факта были получены А. В. Кщбяковым в 1933 г. в опытах, в которых он пропускал через сосуды верхнего шейного симпатического узла кошки раствор Рингера — Локка и обнаружил при раздражении преганглионарных симпатических волокон в растворе, оттекающем от узла, адреналиноподобное вещество. В дальнейшем было показано, что возбуждающим медиатором в синапсах преганглионарных волокон является ацетилхолин. Адреналин оказался медиатором, вызывающим; торможение активности нейронов симпатического ганглия. Возможно, что тормозящие волокна, в которых образуется адреналиноподобное вещество, представляют собой постганглионарные волокна, иннервирующие узел и изменяющие его функциональное состояние. Особенностью действия ацетилхолина в синапсах ганглиев является, то, что оно не прекращается после отравления.узла атропином, но исчезают после отравления никотином. На этом основании считают, что существует два вида структур, чувствительных к ацетилхолину;.одни из них — М-холинорецепторы — теряют чувствительность к ацетилхолину под влиянием атропина, другие — Н-холинорецепторы — под влиянием никотина и других веществ, называемых ганглиоблокаторами (гексоний и др.). В области концевых разветвлений симпатических нервных волокон имеются расширения — варикозы, в которых находятся пузырьки — везикулы, подобные имеющимся в синапсах. Толщина этих расширений концевых нервных волокон — до 2 мкм, длина — 0,5—3 мкм. Таких варикозов может быть 15—30 на протяжении 100 мкм. В варикозах содержится в 20—100 раз больше норадреналина, чем в остальных участках постгангли- онарного волокна. В расширенной части концевых разветвлений, а не только в синапсах симпатических нервных волокон возможно высвобождение медиатора, действующего на иннервированную ими ткань. В зависимости от того какой медиатор выделяется окончаниями аксонов вегетативных нейронов, предложено разделять нейроны на холинергические и адренергические. Холинергическими являются эфферентные нейроны интрамуральных парасимпатических ганглиев и эфферентные нейроны парасимпатических центров среднего, продолговатого и спинного мозга, а также эфферентные нейроны симпатических центров спинного мозга и те эфферентные нейроны периферических симпатических ганглиев, которые иннервируют потовые железы и обеспечивают расширение сосудов работающих мышц. Окончания аксонов этих нейронов выделяют ацетилхолин. Адренергическими являются все остальные эфферентные нейроны симпатических ганглиев. В окончаниях аксонов и в контактах, образованных этими аксонами с гладкомышечными клетками и другими структурами, выделяется норадреналин. Освобождающийся в терминалях аксонов медиатор — ацетилхолин. йли норадреналин взаимодействует со специфическим белком постсинаптической мембраны, образующим комплексное соединение с медиатором. Белок, с которым взаимодействует ацетилхолин, получил название холинорецептора, а белок, взаимодействующий с адреналином'или норадреналином, назван адренорецеп- тором. Соединение медиатора с соответствующим рецепторным веществом является начальной реакцией в цепи химических превращений, возникающих в клетке под влиянием приходящих к ней нервных импульсов. Имеется два основных вида адренорецепторов, с которыми взаимодействует как адреналин, так и норадреналин: а- и $-адренорецепторы. Их существование установлено путем применения некоторых фармакологических препаратов, действующих избирательно на тот или другой вид адренорецепторов. В ряде органов находится оба вида адренорецепторов, которые могут вызывать либо разные, либо одинаковые реакции, или же имеется только один из адренорецепторов. В кровеносных сосудах имеются и а- и p-адренорецепторы. Показано, что соединение симпатического медиатора с а- адренорецепторами в артериальной стенке вызывает сужение артериол, а соединение с р-адренорецепхорами приводит к расширению артериол. В кишечнике также имеются и а- и (3-адренорецепторы; воздействие и на те и на другие тормозит сокращение гладкой мускулатуры. В сердце и бронхах нет а-адренорецепторов и здесь норадрена- лин и адреналин взаимодействуют только, с p-адренорецепторами. В результате этого происходит усиление сердечных сокращений и расширение бронхов. В механизме действия норадреналина и адреналина придают значение тому недавно открытому факту, что норадреналин и адреналин активируют энзим, находящийся в мембране мышечных клеток,— аденилциклазу. Этот энзим в присутствии ионов магния катализирует образование в клетке из АТФ циклического 3,5-аденозинмонофосфата. Это соединение — цАМФ вызывает ряд физиологических эффектов, в частности активирует некоторые энзимы энергетического обмена и стимулирует сердечную деятельность. Кроме ацетилхолина и норадреналина,' в вегетативной нервной системе найдены и другие медиаторы. В окончаниях симпатических нервных волокон обнаружен дофамин, выделение которого в синаптическую щель происходит под влиянием приходящих нервных импульсов. Полагают, что дофамин вступает во взаимодействие с а-адренорецеп- торами, расположенными на самих пресинаптических окончаниях, и тем самым тормозит выделение норадреналина. Полагают, что на гладкую мускулатуру кишечника, матки, а возможно, и кровеносных сосудов может действовать серотонин, эффект которого напоминает действие медиатора ацетилхолина, но сохраняется после блокады М-холинорецепторов. В желудке и кишечнике обнаружены интрамуральные эфферентные нейроны, возбуждение которых тормозит активность гладкой мускулатуры. Это торможение осуществляется путем выделения окончаниями аксонов этих нейронов пуринового нукле- отида аденозинтрифосфорной кислоты (АТФ). Медиаторный эффект принадлежит, по-видимому, самой АТФ. Указанные эфферентные нейроны получили название пури- нергических. Предполагают, что медиатором может быть и гистамин, так как в некоторых тканях обнаружены специфические Н|- и Нй-гистаминорецепторы. Гистамин является биологически активным веществом широкого спектра действия. Выявлено, что широко распространенный в синапсах ЦНС тормозной медиатор гамма-аминомасляная кислота — ГАМК тормозит проведение возбуждения в звездчатом ганглии, но облегчает передачу возбуждения в верхнем шейном, нижнем брыжеечном и в ганглиях солнечного сплетения. После перерезки и перерождения вегетативных нервов чувствительность денерви- рованных органов к соответствующим медиаторам возрастает. Если десимпатизировать любой орган, иннервированный симпатическими нервными волокнами (сердце, желудок, кишечник, сосуды, радужную оболочку глаза и др.), то он приобретает повышенную чувствительность к адреналину и норадреналин'у. Точно так же, если произвести парасимпатическую денервацию органа, он может приобрести повышенную чувствительность к ацетилхолину. Имеется ряд механизмов этой повышенной чувствительности денерви- рованных тканей. Среди них следует указать на возрастание числа рецепторов на постсинаптической мембране, снижение активности или содержания в тканях фермента, расщепляющего адреналин (моноаминооксидаза) или расщепляющего ацетилхолин (ацетилхолинэстераза) и др. ВЕГЕТАТИВНАЯ ИННЕРВАЦИЯ ТКАНЕЙ И ОРГАНОВ ЗНАЧЕНИЕ ВЕГЕТАТИВНОЙ ИННЕРВАЦИИ Роль вегетативной нервной системы заключается в регуляции обмена веществ, возбудимости и автоматии периферических органов, а также самой ЦНС. Вегетативная нервная система регулирует и изменяет физиологическое состояние тканей и органов, приспосабливая их ^ к текущей деятельности целостного организма и условиям окружа юшей среды. В зависимости от условий функционирования органов вегетативная нервная систем; оказывает на них корригирующее и пусковое влияние. Если орган обладает автоматие! и непрерывно функционирует или «запущен в работу», а импульсы, приходящие п< симпатическим или парасимпатическим нервам, только усиливают или ослабляют ег< деятельность, в таком случае говорят о корригирующем влиянии. Если же работ; органа не является постоянной, а возбуждается импульсами, поступающими по симпати ческим или парасимпатическим нервам, в этом случае говорят о пусковом влиянии веге тативной нервной системы. Пусковые влияния нередко дополняются корригирующими ВЛИЯНИЕ СИМПАТИЧЕСКОЙ И ПАРАСИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ НА ФУНКЦИЮ ОРГАНОВ В большинстве органов, иннервируемых вегетативной нервной системой, раздраже ние симпатических и парасимпатических волокон вызывает противоположный эффект Так; сильное раздражение блуждающего нерва вызывает уменьшение ритма и силь сердечных сокращений,'раздражение симпатического нерва увеличивает ритм и сил; сердечных сокращений; парасимпатические влияния расширяют сосуды языка, слюнны: желез, половых органов, симпатические -— суживают эти сосуды; парасимпатически' нервы суживают зрачок, симпатические — расширяют; парасимпатические влияни: суживают бронхи, симпатические — расширяют; блуждающий нерв стимулирует работ; желудочных желез, симпатический — тормозит; парасимпатические нервы вызываю расслабление сфинктеров мочевого пузыря и сокращение его мускулатуры, симпатиче ские сокращают сфицктер. и расслабляют мускулатуру и т. д. Эти факты позволили выдвинуть гипотезу об «антагонизме» симпатического и парасимпати ческого отделов вегетативной нервной системы. Согласно этой гипотезе, оба отдела управляю функцией органа, действуя в противоположном направлении (подобно двум вожжам), В норм наблюдается «равновесие» между тонусами двух отделов вегетативной нервной системы, т. < уравновешивание влияний симпатического отдела нервной системы влияниями парасимпатическог отдела. При преобладании тонуса одной системы тонус другой уменьшается. Постоянное повышенн тонуса симпатического или парасимпатического отдела приводит к появлению различных рас стройств — «симпатикотоний» и «ваготоний». Предпринимались попытки лечить «симпатикотонии и «ваготонии» хирургической перерезкой соответствующих нервов, Однако, подобные воздействи не давали стойкого эффекта, а иногда приводили и к ухудшению состояния. Многочисленные экспериментальные исследования показали, что между двумя отделам вегетативной нервной системы существует не только антагонизм, но и синергизм. Повышени тонуса одного из отделов неизбежно вызывает процессы, приводящие к повышению тонуса другогс Полагали, что нормальная работа органов может протекать лишь в случае «уравиовешива ния» симпатических влияний парасимпатическими. Однако ряд факторов противоречил и этом; допущению: секреция слюны возбуждается как симпатическими, так и парасимпатическим! нервами. Ряд органов и тканей не имеет парасимпатической иннервации, а снабжается тольк< волокнами симпатической нервной системы. К:ним относятся сосуды кожи, некоторые сосудь брюшной полости, мозговой слой надпочечника, матка, скелетные мышцы, органы чувств i сама ЦНС. В опытах, проведенных Л. А. Орбели и А. Г. Гинецинским, . регистрировала сокращения икроножной мышцы лягушки при продолжительном ритмическом раздра женин иннервирующих ее VIII и IX передних корешков спинного мозга. По мере утомле ния мышцы амплитуда сокращений постепенно уменьшалась. Когда сокращения стано вились очень малыми, к продолжающемуся ритмическому раздражению передние корешков присоединяли непродолжительное раздражение симпатического погранцчног( ствола в области тех ганглиев, которые снабжают симпатической иннервацией икронож ную мышцу. Амплитуда сокращений, которыми мышца реагировала на ритмически! Рис. 107. Влияние раздражения симпатических волокон на кривую мышечного утомления изолированной икроножной мышцы лягушки (по Орбели — Гинецинскому). Сокращения утомляемой мышцы вызываются ритмическим (30 раз в минуту) раздражением двигательных нервных волокон. Моменты раздражения симпатического нерва отмечены поднятием сигнальной линии. раздражения двигательных, корешков, начинала постепенно расти и иногда достигала первоначальной (рис. 107), т. е. утомление мышцы устранялось. Было выявлено, что симпатическая нервная система оказывает влияние на органы чувств. Импульсы, идущие по симпатическим путям, действуют также на ЦНС, в частности на рефлекторную функцию продолговатого и среднего мозга, а также на условно- рефлекторную деятельность коры больших полушарий. По данным некоторых-авторов, после удаления верхних шейных симпатических узлов у собаки наблюдаются нарушения условнорефлекторной деятельности. Основываясь на этих фактах, Л. А. Орбелй высказал положение об универсальной адаптационно-трофической функции симпатической нервной системы. Согласнд этой точке зрения, симпатическая система регулирует обмен веществ, трофику и возбудимость всех органов и тканей тела, обеспечивая адаптацию организма к текущим условиям деятельности. Если симпатический отдел играет универсальную адаптационно-трофическую роль, го остается неясным физиологическое значение парасимпатического отдела. Ряд фактов свидетельствует о том, что симпатический отдел вегетативной нервной системы 1ктивирует процессы, связанные с расходом энергии, а парасимпатический — с ее накоплением ) организме. Появилась точка зрения, что «антагонизм» между этими двумя отделами проявляется 1менно в том, что симпатические влияния активируют процессы, связанные с деятельностью организма, а парасимпатические влияния способствуют восстановлению тех ресурсов, которые ютрачены при этой деятельности. Однако известно, что ряд'органов, иннервируемых симпатическими нервами (скелетные мышцы, органы чувств, сама ЦНС), весьма активно функционируют 1ри напряжении сил, однако не имеют парасимпатической иннервации. А именно эти органы г первую очередь нуждаются в восстановлении своих ресурсов, потраченных при напряженной деятельности. • Как видно из сказанного, ни одна из приведенных точек зрения не смогла объяснить противо- >ечия и ответить на вопрос о взаимоотношениях симпатического и парасимпатического отделов. В последнее время было п<жазано, что взаимоотношение двух отделов вегетативной юрвной системы не может быть выражено понятиями «антагонизм» и «синергизму. Каждая из систем выполняет свою собственную функцию в организме. До последнего времени роль парасимпатического отдела, как правило, изучалась путем :верхпорогового раздражения электрическим током парасимпатических нервов. Однако подобный [рием исследования мог приводить к ошибочным выводам. Известно/что парасимпатические нервы :остоят из тысяч нервных волокон, каждое из которых имеет свой собственный «код» для передачи юобщений. Индивидуальность этого «кода» проявляется в определённой частоте импульсов данного юл окна, в характере группирования импульсов в пачки, в количестве импульсов, составляющих [ачку, в величине интервалов между пачками, в сочетании интервалов между импульсами в соседних юлокнах, в характере сочетания во времени возбужденных и невозбужденных волокон и т. д. 02000201020100010101003001020202000100 Исследователи же пользовались, как правило, лишь сильными одновременными раздраже- шями всех волокон/сразу, что равносильно дому, как, если бы вместо мелодии, извлекаемой из фортепиано музыкантом-пианистом, нанести удар бревном одновременно по всем клавишам. Тонкие приемы исследования выявили, что влияние парасимпатических волокон н; деятельность органов неоднозначно. Волокна парасимпатического отдела действую' на изучаемые органы и ткани не непосредственно. В стенке иннервируемого орган; имеются эфферентные нейроны, на которых оканчиваются преганглионарные волокна Интрамуральные эфферентные нейроны представляют собой общий конечный путь дл! импульсов, поступающих из ЦНС по преганглионарным волокнам парасимпатически: нервов, и для импульсов, приходящих к ним от внутриорганн*?1х рецепторов, сформиро ванных интрамуральными афферентными нейронами, представляющими собой перво< звено интрамуральной периферической рефлекторной дуги. Итак, интрамуральные нервные ганглии не только передаточные станции дл9 импульсов, поступающих из ЦНС по преганглионарным волокнам парасимпатически: нервов. Нейроны, входящие в эти ганглии, их отростки, синапсы и окончания формирую' внутриорганные рефлекторные структуры, регулирующие работу органа путем внутри органных периферических рефлексов. Импульсы, приходящие к органу по преганглио нарным волокнам парасимпатических нервов, вступают во взаимодействие с импуль сами, осуществляющими процессы внутриорганной рефлекторной регуляции. Характе] ответной реакции органа определяется результатом указанного взаимодействия. Поэ тому эффект раздражения преганглионарных волокон не бывает однозначным. (Ка) было сказано выше, однозначные эффекты возникают лишь в результате нефизиологи ческих методов исследования — сильнейшего одновременного раздражения всех пре ганглионарных волокон). На органы, в которых выявлено существование интрамуральных рефлекторные механизмов регуляции, преганглионарные парасимпатические волокна могут оказыват! (в зависимости от функционального состояния иннервируемого органа) как возбуждаю щее, так и тормозящее влияние. ' ■ ' Противоположные влияния парасимпатических волокон отнюдь не являются «парадоксальными». Это естественное проявление разнонаправленных воздействий необходимых для обеспечения нормальной функции органов и тканей. Парасимпатиче с кий отдел представляет собой систему, способную осуществлять текущую регуляцик физиологических процессов и обеспечивать в полном объеме поддержание постоянства внутренней среды организма. Она располагает для этого огромными возможностями Достаточно указать, например, на существование в кишечнике трех нервных сплетений содержащих большое количество ганглиев. Количество интрамуральных нейронов, при ходящихся на 1 см 2 поверхности кишечника, может достигать 20 ООО. Структура интрамуральных ганглиев напоминает нервные центры, вынесенные ш периферию. Каждый нейрон окружен большим количеством клеток нейроглии. Имеются структуры, избирательно пропускающие к нейрону из крови лишь определенные вещества, напоминающие по своей функции гематоэнцефалический барьер. Таким образом, нейроны ганглия, подобно нейронам мозта, защищены от непосредственного воздей ствия веществ, циркулирующих в крови. Среди интрамуральных эфферентных нейроноЈ имеются не только холинергические, но и адренергические, а также пуринергические серотонинергические, дофаминергические и, по-видимому, гистаминергические, пепти- дергические-nГАМК-ергические. Все это создает возможность для большого диапазона регуляторных воздействий. Как было сказано выше, импульсы, приходящие к органу по преганглионарным волокнам парасимпатических нервов, вступают во взаимодействие с импульсами, осуществляющими процессы внутриорганной рефлекторной регуляции и в зависимости от текущего состояния физиологических процессов, протекающих в данном органе или системе, могут включать или выключать, усиливать или ослаблять ту или иную функцию органа, осуществляя многообразные регуляторные влияния, необходимые для поддержания нормальной текущей деятельности и гомеостаза. ' В отличие от этого симпатический отдел вегетативной нервной системы при разной силе раздражения оказывает на органы однотипные влияния. Волокна симпатической системы^ подходящие к органу —- это постганглионарные волокна. Они оканчиваются непосредственно на рабочих структурах органов и ■тканей. Импульсы, приходящие по этим волокнам, не вступают во взаимодействие с аппаратами внутрйорганной рефлек- горной регуляции, а оказывают на работу органа прямое и однотипное влияние. Это обусловлено тем, что симпатический отдел играет особую биологическую роль, заключающуюся-в мобилизации сил и резервов организма для преодоления трудностей, решения гложнейших задач, возникающих при активном взаимодействии организма и окружающей среды. Из сказанного понятно, что парасимпатический отдел вегетативной нервной системы —- это система текущей регуляции физиологических процессов, обеспечивающая гомеостаз. В отличие от этого симпатический отдел — это система тревоги, система «защиты», система мобилизации резервов, необходима^ для активного взаимодействия организма со средой. Такая мобилизация требует генерализованного включения в реакцию многих органов и структур. По-видимому, именно поэтому ганглии симпатического отдела (пар.авертебральные и превертебральные) находятся на большом расстоянии от иннервируемых органов и тканей и обладают большими возможностями умножения (мультипликации) импульсов, что обеспечивает генерализованное воздействие на многие структуры. Генерализованное воздействие почти на все структуры организма возникает и при выбросе в кровь адреналина из хромаффинной ткани. (Адреналин поэтому представляет собой «жидкую симпатическую нервную систему»). Симпатический отдел вегетативной нервной системы активирует деятельность мозга, мобилизует защитные реакции: процессы терморегуляции, иммунные реакции, механизмы свертывания крови, барьерные механизмы. Ее возбуждение—непременное условие состояния напряжения и стенических эмоций. Возбуждение симпатической системы является начальным звеном включения цепи гормональных реакций, характерных для «стресса». В отличие от парасимпатического отдела, обеспечивающего'поддержание гомеостаза, симпатический нередко его изменяет. Возбуждение его приводит к повышению артериального давления, опустошению кровяных депо, выбросу в кровь больших количеств глюкозы и жирных кислот, активации энергетических процессов, угнетению функций желудочно-кишечного тракта, мочеобра- зования, выделения мочи и т. д. Жизнь организмов в естественных биологических условиях — непрерывней борьба за существование, в которой побеждает наиболее приспособленный, т. е. наиболее сообразительный, сильный, ловкий, быстрый, неутомимый. У высших организмов в процессе эволюции появилась жизненная необходимость в создании инструмента, максимально мобилизующего двигательную и интеллектуальную активность, запускающего в действие все ресурсы, все резервы организма. Таким инструментом стал симпатический отдел вегетативной нервной системы. Этот отдел нередко дестабилизирует физиологические процессы, обеспечивая максимальное напряжение функций всех тех органов и систем, которые необходимы для огромных усилий, для гигантской мобилизаций интеллектуальных, энергетических ресурсов, для небывалой по мощности и масштабам мышечной деятельности,' для спасения организма путем борьбы или бегства. Из сказанного ясно, что симпатический отдел нередко нарушает постоянство внутренней среды. Задачу восстановить и сохранить постоянство внутренней среды при любых нарушениях и сдвигах, вызванных возбуждением симпатического отдела, падает на долю парасимпатического отдела. В этом смысле деятельность двух отделов может проявляться иногда как антагонизм. Но это не значит, что функции органов и тканей управляются только антагонистическими влияниями. Парасимпатические нервные волокна в ряде случаев могут как стимулировать, так и тормозить функцию регулируемых ими органов, обеспечивая все. процессы текущей-регуляции, необходимые для сохранения гомеостаза. В последнее время показано,, что выделяемый окончаниями парасимпатической системы ацетилхолин может тормозить секрецию норадреналина окончаниями симпатической нервной системы и, кроме того, понижать чувствительность адренорецепторов к действию катехоламинов. Таким образом, парасимпатическая система может играть роль и регулятора (модулятора) симпатических влияний, являясь .своеобразным «антистрессорным» фактором. Задача парасимпатического отдела вегетативной нервной системы — непрерывно корригировать сдвиги, вызванные влиянием симпатического отдела, восстанавливать и сохранять гомеостаз. ВЕГЕТАТИВНЫЕ РЕФЛЕКСЫ И ЦЕНТРЫ РЕГУЛЯЦИИ ВЕГЕТАТИВНЫХ ФУНКЦИЙ ВЕГЕТАТИВНЫЕ РЕФЛЕКСЫ Нейроны вегетативной нервной системы участвуют в осуществлении многих рефлекторных реакций, называемых вегетативными рефлексами. Последние могут быть вызваны раздражением как экстерорецепторов, так и интерорецепторов. При вегетативных рефлексах импульсы передаются из ЦНС к периферическим органам по симпатическим или парасимпатическим нервам. Число вегетативных рефлексов очень велико. В медицинской практике имеют большое значение висцеро-висцеральные, висцеро-дермальные и дермовисцеральные рефлексы. Висцеро-висцеральные рефлексы— реакции, которые вызываются раздражением рецепторов, расположенных во внутренних органах, и заканчиваются изменением деятельности также внутренних органов. К числу висцеро-висцеральных рефлексов относятся рефлекторные изменения сердечной деятельности, тонуса соСудов, кровенаполнения селезенки в результате повышения или понижения давления в аорте, |