Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
 Скачать 7.39 Mb. Скачать 7.39 Mb.
|

^Нейронная организация. Несмотря на значительную функциональную специализацию различных областей коры, их нейронная структура имеет много общего. Стереотипность нейронной организации коры мозга проявляется в упорядоченном расположении нервных клеток и волокон (рис. 96). Нервные элементы коры больших полушарий ориентированы послойно, образуя 6 эсновных слоев. Слой I, наиболее поверхностный, плексиформный или молекулярный, содержит незначительное число нервных клеток. Он образован главным образом сплетением' нервных волокон. Во II слое, называемом наружным зернистым, плотно расположены мелкие (4— 3 мкм) нейроны, тела которых имеют овальную, треугольную или многоугольную форму (клетки-зерна). , , ВIII слое расположены пирамидные нейроны разных размеров. Слой IV, называемый внутренним зернистым слоем, содержит подобно наружному зернистому слою скопление мелких нейронов. ■ Слой V состоит из гигантских пирамидных клеток, или клеток Беца. Вверх от них зтходят длинные дендритные отростки, достигающие поверхностных слоев,— так назы- заемые апикальные дендриты. Базальные дендриты пирамидных.клеток занимают боковое положение. Аксоны крупных пирамидных нейронов проецируются к различным ядрам Уловного и спинного мозга. Самые длинные из них образуют пирамидный тракт, достига- ощий каудальных сегментов/спинного мозга. Слой VI мультиформный, содержит нейроны веретенообразной- и треугольной формы. Главными эфферентными нейронами коры являются большие пирамидные клетки V слоя, аксоны которых покидают ее пределы. Более мелкие, короткоаксонные нейроны обеспечивают внутрикорковые связи и выполняют роль возбуждающих и тормозных вставочных нейронов. Характерную организацию имеют также приходящие в кору афферентные волокна. Главный афферентный вход в кору больших полушарий образован таламокорти- кальными проекциями. Таламические волокна образуют в коре два основных типа окончаний. v. Волокна, приходящие в кору от нейронов специфических (проекционных) ядер таламуса (зрительных, слуховых), доходят до наружного зернистого слоя и оканчиваются густыми разветвлениями внутри III и IV слоев. Добавочные их разветвления обнаруживаются и в VI слое. Благодаря густому ветвлению окончаний проекционных волокон вокруг клеток III и IV слоя обеспечивается возможность эффективно доставлять, афферентную информацию в топографически определенные зоны и подвергать ее локальной обработке на уровне: вставочных нейронов, .' Другой тип окончаний: таламокорти- кальных проекций образуется аксонами нейронов неспецифических ядер таламуса. Они распространяются по широким областям коры и дают коллатерали, проходящие через все кортикальные слои, достигая и самого поверхностного слоя. Таким образом обеспечивается, универсальный настроенный меха низм, влияющий на уровень возбудимости пирамидных нейронов, и интеграция корково* активности в целом. В настоящее время обнаружено значительное многообразие типов неспецифически тдламокортикальных волокон. Во многих областях коры обнаруживается конвергенция проекций от несколькю таламических ядер. ' 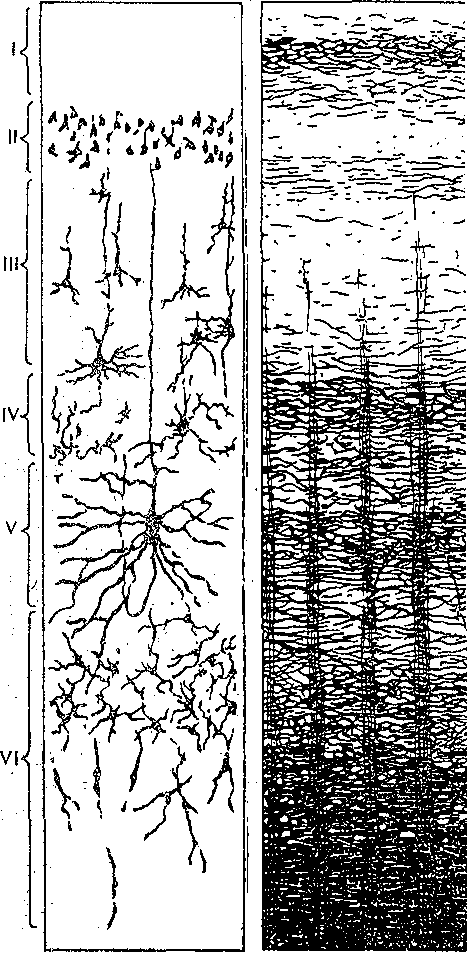 Рис. 96. Расположение нейронов и нервных волокон в коре большого мозга. l—V-I —- номера слоев коры (объяснение в тексте) Несмотря на. известную'общность нейронной организации разных отделов коры более детальный анализ показывает определенные различия, проявляющиеся в ходе волокон, количестве и размерах нейронов, ветвлении дендритов. Эти различия обусловлены неодинаковой функциональной специализацией разных областей коры. На основании цитоархитектонических признаков была построена карта коры, в которой выделень различные корковые поля. На карте в коре больших полушарий человека выделено 1 [ .областей, включающих 52 поля. Функции древне^ и старой коры 'Для обозначения всех корковых образований, не относящихся к новой коре,; древнюю и старую кору часто объединяют. На основании морфологических данных выдвинута точка зрения на древнюю и старую кору как на особую систему переднего мозга,, названную обонятельным, или висцеральным, мозгом. Согласно этой точке зрения, обонятельный мозг, помимо ф.унк^ ций, связанных с обонянием, ведает реакциями настораживания и внимания,, принимает участие в регуляции вегетативных функций. Эта система играет также важную роль в осуществлении инстинктивного поведения (пищевого, полового, оборонительного) и в формировании эмоций. Помимо обонятельной афферентации в старую кору поступает также импульсация от других сенсорных путей, причем наблюдается значительная конвергенция различных афферентных влияний на одних и тех же нервных элементах, .■ . _ Раздражение структур старой коры оказывает влияние на сердечно-сосудистую систему и дыхание. Повреждения в области старой коры могут вызвать гиперсексуальность, изменять эмоциональное поведение. Имеются зоны, раздражение которых приводит к реакции насыщения и удовольствия. При электрическом раздражении миндалины наблюдаются эффекты, связанные с деятельностью пищеварительного тракта: облизывание,.жевание, глотание, саливация, изменения желудочной секреций, перистальтики ;кишечника. Раздражение миндалины влияет также на деятельность и других внутренних органов:; почек, мочевого пузыря, матки. ; Таким образом, существует тесная связь структур.старой коры с вегетативной нервной системой, с процессами, направленными на регуляцию внутренней среды организма. Эта деятельность старой коры протекает при участии гипоталамуса. Древняя и старая кора вместе с гипоталамусом и лимбической областью среднего, мозга составляют так называемую лимбическую систему, обеспечивающую гомеостаз, самосохранение и сохранение вида. Имеются данные, что некоторые области старой коры имеют важное значение,в процессах памяти. При удалении .миндалевидного комплекса нарушаются отставленные рефлексы. У больных с нарушениями памяти при патоморфологических исследованиях нередко обнаруживаются признаки дегенеративных явлений в гиппокампе. . Электрофизиологические исследования активности нейронов гиппокампа показали, что их ответы на афферентные; раздражения отличаются большой продолжительностью. Большое значение для интенсивности реакции имеет новизна раздражения. Интересной особенностью синаптического аппарата гиппокампа оказалась способность чрезвычайно длительное время удерживать следы предшествующего раздражения. Так, посттетани- ческая потенциация в ответ на короткое тетаническое раздражение в синапсах пирамидных нейронов гиппокампа может продолжаться недели и.месяцы, тогда как в спинном мозге лишь немногие минуты: Функции новой коры Исследования эффектов удаления различных участков коры, а также последствий их поражения патологическими процессами, прямого электрического раздражения коры с регистрацией электрической активности показали, что различные поля коры больших полушарий тесно связаны с определенными функциями. Так, ;при применении электрической стимуляции была выявлена возможность получения двигательных реакций при раздражении определенных корковых зон, что привело к представлению о существовании моторной коры. Определение зон, в которые проецируются специфические .афферентные системы, позволило выделить сенсорные области коры, представляющие топически организованные проекции различных периферических рецепторных полей. Выявлены обладающие ассоциативной функцией зоны, в которые поступает информация от афферен- то в различных модальностей. Имеются большие корковые поля; функция которых менее определенна. Так, значительная часть лобных долей (поля 9 и,12) не отвечает на электрическое раздражение й может быть удалена (особенно в правом полушарии) без заметных'последствий. Однако двустороннее удаление большей части полей 9 и 12 ведет к значительным психическим нарушениям. Таким образом, зоны, воспринимают^ сенсорные сигналы или посылающие эфферентные импульсы к нижележащим Центрам, имеют наиболее фиксированную локализацию в' коре, в то время как ассоциативные поля связаны главным Образом со'сложными Процессами высшей нервной деятельности и функционируют как целое. Сенсорные зоны коры больших полушарий Афферентные:импульсы, направляющиеся в кору больших полушарий, переключаются на клетках ядер таламуса и оттуда проецируются в определенные корковые поля. В каждом полушарии. выделяются первичные зоны .представительства соматической (кожной и мышечно-суставной) и висцеральной чувствительности. Эти зоны обозначаются как I и II соматосенсорные зоны коры. Первая соматосенсорная зона расположена в задней центральной извилине, и имеет значительно большую площадь, чем вторая. К ней поступают волокна от заднего вентрального ядра таламуса. Особенно большую поверхность занимает представительство рецепторов кисти рук, голосового аппарата и лица, значительно меньшую •• туловища, бедра и голени. Эти различия отражают различия в количестве реценторных образований, имеющихся в коже туловища и наиболее чувствительных участков тела — губах, языке, подушечках Пальцев. . Вентральнее I соматосенсорной зоны, в латеральной (сйльвйевой) борозде, обнаружена // соматосенсорная зона, куда также поступают волокна от клеток заднего вентрального ядра таламуса. Постцентральнан область коры часто подвергалась прямому электрическому раздражению у пациентов во время хирургических операций. Результатом такого раздражения были ощущения давления, прикосновения или тепла, ощущаемые на-противоположной половине тела. ... Удаление участков соматосенсорной области приводит к потере тонкой градации чувствительности той Части тела-, которая представлена в удаленном участке коры. Кроме того, появляется определенная неловкость и неаккуратность при движениях данной части тела. Таким образом,, главная функция соматосенсорной области заключается в интеграции и критической оценке той информации, которая поступает из специфических ядер таламуса. Здесь происходят оценка1 относительной интенсивности ощущений, определение пространственных взаимоотношений раздражаемых участков тела, выявление сходства и. различия ощущаемых раздражений. Другой четкой первичной проекционной Областью коры является внутренняя поверхность затылочной коры в области шпорной борозды. В эту область поступают аксоны клеток наружного коленчатого тела, доставляющие в кору зрительную информацию. В I зрительной области (поле 17) обнаруживается топографически организованное представительство сетчатки. У млекопитающих в связи с бинокулярным, зрением первичная зрительная область каждого полушария получает проекции сетчаток обоих глаз. При этом в каждое полушарие проецируются одноименные половины сетчаток (в левое — обе правые их половины, а в правое — обе левые). •...'' При электрическом раздражении 17-го поля человек испытывает световые ощущения. Поля 18 и 1,9 связаны с ассоциацией зрительной и других Видов чувствительности. Здесь зрительные, тактильные и отчасти слуховые воздействия Подвергаются синтезу, обеспечивая более полное зрительное ощущение. Повреждения полей 18 и 19 приводят к нарушению, зрительной оценки, так что, например, написанные или напечатанные слова не воспринимаются. Раздражение 19-го поля вызывает зрительные галлюцинации, движения глаз. - ' Слуховая зона коры (поля 41и 42) в основном расположена в латеральной борозде. Только небольшая часть этой зоны видна на верхнем крае Височной доли. В данной области коры звуковые сигналы, попадающие в улитку, воспринимаются как звуки, вар*>т ирующие по тону, громкости и качеству. Сенсорная слуховая область отличается богатством клеточного и волоконного состава, имеет хорошо выраженный слой крупных пирамидных нейронов. Типичным для слуховой коры является также хорошо выраженный слой звездчатых клеток. В слуховую область, кроме слуховых путей, проецируются также вестибулярные афференты. . Электрическое раздражение слуховой области коры у людей с сохраненным сознанием вызывает субъективные ощущения шума в обоих ушах. Благодаря двустороннему представительству поражение височной доли на одной стороне хотя и вызывает нарушения слуха, но не приводит к глухоте. В слуховой области коры имеется топическое представительство различных частей улитки. Зона, лежащая на периферии первичной слуховой области, образует ассоциативный центр, связанный преимущественно с оценкой значимости звуков. Раздражение этой зоны вызывает слуховые галлюцинации и движения головы в противоположную сторону. Специальные зоны верхнего и среднего височных выпячиваний (конволюций) в левом полушарии связаны, с пониманием речи. При их повреждении теряется способность произносить или понимать произнесенные слова. Моторные зоны коры больших полушарий В коре больших полушарий выделяются зоны, раздражение которых закономерно вызывает четкие, координированные двигательные реакции. У приматов и, человека двигательная область расположена в прецентральной извилине (поля 4 и 6). Кроме того, имеется еще и дополнительная- двигательная область, располагающаяся на медиальной поверхности коры. - Для двигательной области коры характерна строго топическая организация распределения двигательных функций. Слабое электрическое раздражение определенных точек моторной коры вызывает движение определенных мышц противоположной половины тела. Наиболее дискретные движениями с наименьшим порогом вызываются при стимуляции 4-го ноля. \ Здесь, в V слое коры находятся особенно крупные пирамидные клетки — гигантские пирамидные нейроны, превосходящие своими размерами пирамидные нейроны всех остальных корковых полей. Аксоны гигантских пирамидных нейронов образуют наиболее быстро проводящие проекции пирамидного тракта. Эти волокна Достигают каудальных сегментов спинного мозга, а их окончания образуют возбуждающие синапсы непосредственно на мотонейронах передних рогов. Поскольку моносинаптические контакты между окончаниями аксонов пирамидных нейронов и мотонейронами наиболее важны для управления! тонкими движениями, в первую очередь пальцев; после повреждения пирамидного тракта или 4-го поля коры дискретные движения в ответ на стимулы, приложенные к моторной коре, или произвольные команды значительно нарушаются. ■ " . Различные мышцы представлены в строго определенной.последовательности впереди центральной борозды. Как и в случае соматосенсорной коры, мышцы разных частей тела представлены неравномерно. Указанное распределение хорошо совпадает с выраженностью моносинаптических связей между пирамидными клетками коры и мотонейронами. Наибольшее число синапсов окончания пирамидных волокон образуют на мотонейронах, иннервирующих мышцы пальцев, лицевую мускулатуру, язык. Поражение моторной коры вызывает параличи и парезы, особенно заметно проявляющиеся в кистях рук, стопах, мимической мускулатуре, мышцах, связанных с артикуляцией. Аксоны нейронов двигательной коры не только образуют кортико-спинальный тракт, но и направляются к различным ядрам экстрапирамидной системы, к которой относятся эфферентные проекции, идущие к нервным клеткам ряда стволовых структур: красному ядру и ретикулярным ядрам (дающим начало ретикулоспинальному тракту). Таким образом, кроме кортико-спинальных нейронов, в двигательной коре имеются также кортико- рубральные, кортико-ретикулярные и другие эфферентные нейроны. В. двигательную область коры также поступает сенсорная информация, причем у приматов большое значение имеет афферентация от мышц и сосудов. По современным представлениям вход от мышечных афферентов в моторную кору настолько специфичен, что обеспечивает осуществление рефлексов, замыкающихся через пирамидные клетки непосредственно на мотонейронах спинного мозга. Через: систему ассоциативных волокон двигательная область коры связана,со многими зонами противоположного полушария, что обеспечивает поступление зрительных, слуховых и других сенсорных сигналов, имеющих важное значение для управления произвольными движениями. Наиболее развитые и топически организованные входы двигательная кора получает из вентролатерального ядра таламуса, клетки которого моносинаптически и полисинап- тически связаны'с пирамидными нейронами. Электрические явления в коре больших полушарий Коре головного' мозга свойственна постоянная электрическая активность,, являющаяся результатом генерации синаптических потенциалов и импульсных разрядов в отдельных нервных клетках. Генерация в коре электрических^колебаний была обнаружена Р. Катоном и А. Данилевским. Возможность регистрации биопотенциалов непосредственно от поверхности головы животных была показана В. Правдич-Неминским в 1925 г. В 1929 г. Г. Бергер зарегистрировал электрическую активность от поверхности головы человека - электроэнцефалограмму (ЭЭГ). В настоящее время различные параметры ЭЭГ хорошо описаны и играют большую роль в оценке состояния различных областей новой коры (рис. 97). В спокойном состоянии у'человека в большей части коры, больших полушарий регистрируется регулярный ритм с частотой около 8-13 Гц в секунду (альфа-ритм) (рис. 97, II). В! состоянии актив- ч ной деятельности он сменяется частыми (более 13 в секунду) колебаниями небольшой амплитуды (бета-ритм) (рис. 97, I). Во время сна Он сменяется медленными (0,5—3,5 в секунду) колебаниями (дельта-ритм) (рис. 97, III, IV, V). -  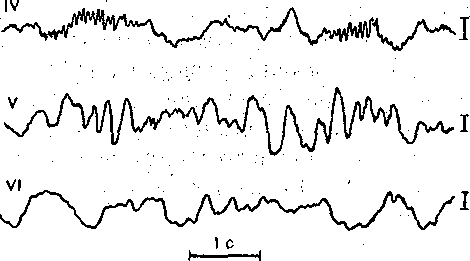 Рис. ■ 97. Типичные электроэнцефалограммы, за писанные, при различных, уровнях сознания.  тмЬ ЧШ* 2Г* Рис. 98. Разряды пирамидного, нейрона коры обезьян (1) при сгибании и разгибании (2) запястья при наличии (а и (5) и отсутствии (в) нагрузки. Аналогичные изменения происходят при активации восходящих путей, проецирующихся в кору. Таким образом, ЭЭГ позволяет судить о функциональном состоянии коры, например о глубине наркоза, о наличии в определенных ее зонах патологических процессов. . ' Для анализа деятельности корковых структур, в особенности у животных,' возможно отведение потенциалов от отдельных нервных клеток. С помощью этого метода удалось охарактеризовать свойства пирамидных и вставочных нейронов, особенности протекания в них синаптического возбуждения и торможения, действия' на их мембрану различных, медиаторов. Большой интерес представляет исследование особенностей активности индивидуальных клеток коры во время выполнения различных функциональных задач. Регистрация активности пирамидных нейронов моторной зоны коры во. время выполнения обезьяной произвольных движений (рис. 98) позволила уточнить характер импульсации этих клеток в связи с двигательной функцией. Кора большого мозга является местом образования условных рефлексов, играющих важнейшую роль в наиболее тонком и точнбм приспособлении организма к условиям окружающей среды. Важной особенностью нейронной организации коры головного мозга является то, что ее нервные клетки образуют особые «элементарные функциональные единицы», представляющие собой колонки, ориентированные перпендикулярно к поверхности. Такие колонки нервных клеток включают все слои коры. - Колончатая организация корковых клеток подтверждается как морфологическими, так и физиологическими исследованиями. Колонкам корковых нейронов присуща тонкая функциональная специализация. Так, в соматосенсорной коре каждая колонка иннерви- рует только одно спинальное моторное ядро и получает строго определенные, топогра-' фически раздельные кожные и лроприоцептивные сигналы с конечности, иннервируемой этим ядром. В пределах колонки можно выделить нейроны с малыми рецептивными полями, отвечающие коротким латентным периодом, и полимодальные нейроны, сложно отвечающие на разнообразные стимулы. Установлено, что нейроны-внутри радикальных колонок имеют топографию частичного перекрытия. Благодаря наличию возвратных коллате- ралей колонки взаимодействуют между собой, Например, по типу латерального торможения. . КООРДИНАЦИЯ ДВЙЖЁЙЙЙ Движение — основная форма активности животных и человека, их взаимодействия с внешней средой. В основе двигательной деятельности лежат процессы координации движений (управления движениями). Они осуществляются -в результате сложного взаимодействия различных отделов ЦНС на основе как врожденных, так и выработанных связей, с участием многих рёцепторных систем (см. гл. 16). Сущностью координации движений является такая пространственная и временная организация процессов возбуждения в мышечном аппарате, Которая обеспечивает выполнение двигательной задачи. Нервные механизмы двигательной деятельности, участие в ней тех или иных отделов ЦНС изучаются в основном в опытах на животных. Однако объектом исследования естественных движений является преимущественно человек. Это. связано с двумя обстоятельствами. Во-первых, человек в зависимости от задачи исследования может воспроизводить любую требуемую форму двигательной деятельности. Во-вторых, движения человека являются проявлением его поведения и трудовой деятельности и поэтому представляют особый интерес как с теоретической точки зрения вследствие их сложности и диф- ференцированности, так и с практической — в связи с их значением для медицины, физиологии труда, космонавтики, эргономики, физиологии спорта. Методы исследования движений человека. Изучение движений человека требует методических приемов, которые были бы безвредными и не нарушали самих двигательных актов. Существует два основных подхода к исследованию движений.'Первый из них— анализ механических параметров движений - траектории, скорости, ускорения, развиваемой силы, второй — непосредственное изучение рйботы Мышц при совершении двигательных актов. '. При анализе механических параметров движения может быть использован принцип циклографии — регистрации последовательных моментов движения путем киносъемки движущегося человека или его фотографирования через равные Промежутки времени на неподвижную пластинку. Если на суставах или точках, соответствующих положению центров тяжести звеньев тела, укреплены метки,.то по такой записи — циклограмме можно восстановить траекторию движения. Существует метод циклогрэмметрии, который дает возможность по циклограммам рассчитать скорость и ускорение, а при учете массы звена — и развиваемую при движении силу. В настоящее время вычислительная техника позволяет автоматизировать подобные исследования. Для регистрации механических параметров движения {механограмм) используют также технику превращения неэлектрических величин в электрические с помощью различных датчиков — магнитных, емкостных, тензометрических и др. Так, с помощью тен- зодатчиков можно непосредственно измерять и' регистрировать силу, прилагаемую к тому или .иному инструменту, или реакции опоры при ходьбе, с помощью резистивных датчиков - регистрировать изменения суставных углов при движении. Электронные дифференциаторы позволяют одновременно с Записью перемещения получать запись скорости и ускорения. . Анализ работы мышц при совершении двигательного акта осуществляется с по-' мощью электромиографии, т. е. регистрации потенциалов действия, возникающих в мышце при ее возбуждении. При электромиографическом изучении движений обычно используют- накожные электроды, укрепляемые, над исследуемой Мышцей. Многоканальный электромиограф дает возможность одновременно записывать электромиограммы нескольких мыщц.' Между .амплитудой зубцов электромиограммы и развиваемой мышцей силой существует примерно Линейная зависимость. Поэтому электромиограммы, в сопоставлении с регистрируемыми синхронно механограммами, позволяют судить о силе сокращения мышц и о распределении их активности в последовательных фазах двигательного акта. ' . : , ...-.. . •• • С помощью описанных выше методов изучены такие сложные естественные двигательные акты, как ходьба, бег, многие рабочие и спортивные движения. Эти методы используются в клинике При исследований нарушений двигательной функции вследствие поражения нервной системы или опорно-двигательного, аппарата. Общие принципы управления движениями. Управление движениями у животных и человека осуществляется нервной системой. По мере филогенетического развития степень.и форма участия .разных отделов мозга в управлении двигательными функциями существенно Менялись. Различны и сами формы двигательной деятельности организмов, ведущих разный образ жизни. У человека двигательные функций достигли наивысшей сложности в связи с переходом к прямостоянию (что осложнило задачу поддержания позы), специализацией передних конечностей на совершение трудовых и других особо тонких движений, использованием двигательного аппарата для коммуникации (речь). В управление движениями человека включены высшие формы Деятельности мозга, связанные с сознанием, что дало основание называть их произвольными. Однако несмотря на сложность и дифференцированность двигательной функции, в ее организации может быть выделен общий фактор, от которого в большой степени зависит иннервацион- ная структура движений. Это - • биомеханические свойства двигательного аппарата, значение которых для координации движений показано Н. А. Бернштейном. Важнейшим биомеханическим свойством опорно-двигательного аппарата позвоночных животных и человека является наличие в скелете большого количества степеней свободы вследствие его многозвенности и двух-, трехосноСти многих суставов. Большое количество степеней свободы обеспечивает чрезвычайное многообразие двигательных возможностей, но при этом делает управление движениями весьма сложной задачей, ■j 149 В каждом движении используются лишь некоторые из степеней свободы, но ЦНС должна постоянно контролировать (ограничивать) все остальные, чтобы обеспечить устойчивость позы. На конечный результат движения влияют не только силы,, развиваемые мышцами, но и силы инерции масс участков тела, вовлекаемых в движение, эластическое сопротивление мышц-антагонистов и связок. Движение смещает различные звенья двигательного аппарата и положения тела, а следовательно, по ходу движения изменяются моменты упомянутых сил. Вследствие изменения суставных углов меняются и моменты мышечных сил. На ход движения влияет также сила тяжести звеньев тела, моменты которой также меняются в процессе движения. В практической деятельности человек вступает во взаимодействие с предметами внешнего мираразличными инструментами, перемещаемыми грузами и т. д., и ему приходится преодолевать силы тяжести, трения, инерции, упругости, возникающие в процессе этого взаимодействия. Немышечные силы вмешиваются в процесс движения и делают необходимым, непрерывное согласование с ними деятельности мышечного аппарата. Необходимо, учитывать также изменение моментов мышечных сил по ходу движения, а также нейтрализовать действие непредвиденных помех движению, которые могут возникать во внешней среде. Все описанное выше делает принципиально необходимым участие в управлении движениями коррекций по ходу движения на основании показаний рецепторов. Таким образом, в управлении движениями можно/выделить два основных механизма. С одной, стороны, при осуществлении любого движения в ЦНС на основе врожденных связей и связей, выработанных в процессе предыдущего двигательного опыта, формируется некоторая пространственно-временная структура возбуждения мышц, соответствующая данной двигательной задаче и исходному положению двигательного аппарата. С другой стороны, важнейшим компонентом управления движениями является внесение по его ходу коррекций в первоначальную структуру мышечного возбуждения. Для характеристики этих двух механизмов используют терминологию кибернетики, называя первый из них программой, второй — коррекциями на основе обратных связей: Участие рецепции в регуляции движений было известно давно. Еще И. М. Сеченов в 1891 г. писал о «согласовании движений с чувствованием». Существенные сведения о роли ироириореценции были получены Фёрстером (1902) и Шеррингтоном (1906). В сенсорном обеспечении движений участвуют, кроме органов зрения и рецепторов мышц, также кожные и суставные рецепторы, вестибулярный аппарат. Относительная роль программ и обратных связей в разных движениях может быть неодинаковой. Так, быстрые движения осуществляются преимущественно на основе программы, медленные, особенно точные — с использованием обратных связей. При обучении новым движениям по мере выработки навыка роль программы возрастает. При .осуществлении даже привычных движений в необычной ситуации, например в невесомости, увеличивается роль обратных связей. Форма участия мышц в осуществлении, двигательных актов весьма многообразна. Анатрмическая классификация мышц (например, сгибатели и разгибатели, синергисты и антагонисты) не всегда соответствует их функциональной роли в движениях. Так, некоторые дву.хсуставные мышцы, в одном суставе осуществляют сгибание, в другом — разгибание. Антагонист может возбуждаться одновременно с агониетом для обеспечения точности движения и его участие помогает выполнять двигательную задачу. В связи с этим, учитывая функциональный аспект мышечной координации, в каждом конкретном двигательном акте целесообразно'выделить основную мышцу (основной двигатель), вспомогательные мышцы (синергисты и другие мышцы, помогающие выполнить двигательную задачу) и стабилизаторы (мышцы, фиксирующие суставы, не участвующие в движении). ' Реализация общих принципов управления движениями может быть рассмотрена на примере конкретных двигательных актов. Ходьба и бег. Ходьба является наиболее обычной формой локомоции человека. Она относится к циклическим двигательным актам, т. е. таким, при которых последовательные фазы движения периодически повторяются. 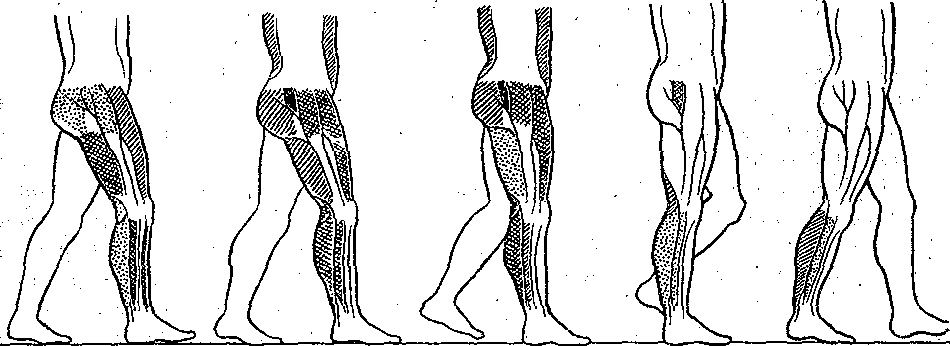 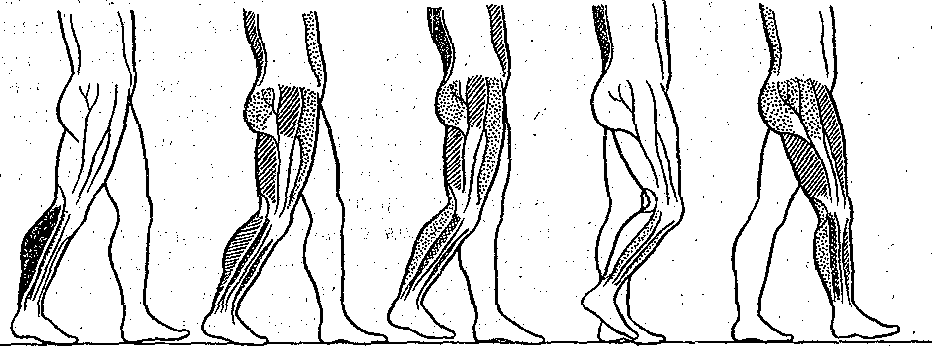 Рис.. 99. Последовательные положения ног при ходьбе. ;.. Показано участие различных мышц в осуществлении движений конечностей. 1, 2 — двухопорный период первого шагз; 6, 7 — двухопорный период второго шага; 3, 4, 5 и 8, 9, !0 —одноопорные периоды. Густота штриховки мышц соответствует силе их сокращений. Для удобства описания и изучения каждый цикл ходьбы подразделяют на периоды: двухопорный период, когда обе ноги стоят на опоре; одноопорный период для правой ноги и переносный для левой, снова двухопорный и затем одноопорный для левой ноги и переносный для правой. Во время опорной фазы происходит перекатывание стопы с пятки на носок. В опорных реакциях ходьбы выделяют передний и задний толчки, первый — при переносе нагрузки ria опорную ногу, второй' — при отталкивании, ноги от опоры. В осуществлении ходьбы участвуют мышцы стопы, голени, бедра и тазового пояса (рис. 99). Сопоставление изменений межзвенных. углов в тазобедренном, коленном и голеностопном суставах с распределением во-времени активности мышц показывает, что кинематика ходьбы является результатом взаимодействия мышечных и немышечных сил. Так, задний толчок, т. е. отталкивание стопы от опоры в результате подошвенного сгибания стопы, осуществляется напряжением задней группы мышц голени, а опускание стопы после соприкосновения пятки с опорой • - под влиянием силы тяжести. Определенные фазы движения в коленном суставе также происходят под влиянием немышечных сил': сгибание в начале фазы переноса осуществляется по инерции в результате заднего толчка, последующее разгибание— сначала вследствие силы тяжести, а затем под влиянием инерции в результате активного движения бедра. . Анализ работы мышц при ходьбе показывает, что в разные фазы шага они сокращаются в разных режимах — концентрическом, т. е. с укорочением (например, мышцы, осуществляющие подошвенное сгибание стопы в опорном периоде), эксцентрическом, г. е: с удлинением (например, сгибатели коленного сустава, тормозящие разгибание в конце переносного периода)^изометрическом, т. е. без изменения длины (мышцы тазобедренного сустава во время переката через пятку). В Последнем случае наблюдается одновременное напряжение мышц-антагонистов, и их функциональная задача при этом заключается в фиксации сустава. В некоторые фазы ходьбы движение имеет баллистический характер, т. е. соответствующая мышца активна лишь в начале перемещения, звена, а за те М движение п родол жается по инерции. ; ' Повторяемость параметров движений в последовательных циклах при ходьбе не абсолютная: они обладают некоторой вариативностью. Наименьшая вариативность у кинематической картины ходьбы, наибольшая — в работе мышц, проявляющаяся в вариативности электромиограмм. Это отражает корригирующую.деятельность ЦНС, которая в каждом шаге вносит в стандартную иннервационную структуру ходьбы поправки, необходимые .для обеспечения относительного постоянства ее кинематики. - Бег отличается от ходьбы тем, что нога, которая находится позади, отталкивается от опоры раньше, чем другая Нога опускается на нее. В результате в беге имеется безопорный период — период полета. В беге благодаря- большим, чем при-ходьбе, скоростям перемещения более значительную роль играют баллистические компоненты движения — перемещение звеньев ноги по инерции. РАБОЧИЕ ДВИЖЕНИЯ Рабочими движениями в широком смысле рл'ова могут'быть названы самые различные целенаправленные движения и действия, совершаемые как в процессе труда, так и в повседневной жизни. У человека основным рабочим органом является рука, причем для выполнения двигательной задачи обычно наиболее важно положение кисти, которая в результате движения должна в определенный момент оказываться в определенном месте пространства. Благодаря большому- количеству степеней свободы верхней конечности кисть может попасть в нужную точку по разным траекториям и при различных соотношениях углов в плечевом, локтевом и лучезаиястном суставах. Это многообразйе'возможностей позволяет выполнять двигательную задачу начиная движение, из различных исходных поз. Электромиографическое исследование ряда рабочих движений показало сложную картину работы мышц, однако в этой картине часто можно выделить устойчивые -сочетания активности нескольких мыщц, используемые в различных движениях. Это так называемые синергии, основанные на врожденных или выработанных в процессе двигательного опыта связях, которые, являясь устойчивыми компонентами движений, упрощают управление сложными двигательными актами. .'.' ' ■ При совершении одного и того же, даже простого движения организация мышечной деятельности в сильной степени зависит от вмешательства немышечных сил, в частности внешних по отношению к человеку. Так, при ударе молотком, когда к массе предплечья добавляется масса молотка и, следовательно, возрастает роль инерции, разгибание предплечья совершается по типу баллистического движения — мышцы-разгибатели активны только в Начале разгибания, которое дальше совершается по инерции, а в,конце притормаживается мышцами-антагонистами. Аналогичное по кинематике движение при. работе напильником, когда основная внешняя сила — трение, совершается путем Непрерывной активности мышцы на протяжении всего разгибания (рис. 100). Если в первом из этих двух движений преобладает роль программы, то во втором велика роль обратных связей. . ПОЗА У млекопитающих животных и человека поддержание позы обеспечивается тем же механизмом сокращения мышц, что и движение. Отличие заключается в том, что при «позной» деятельности мышц сила их сокращения обычно невелика, режим близок к 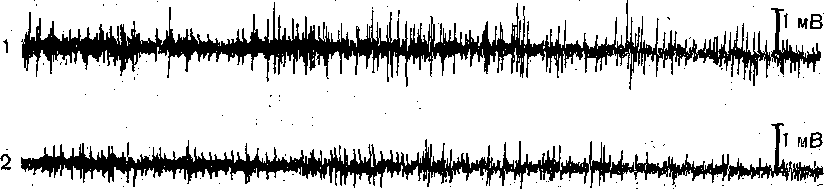 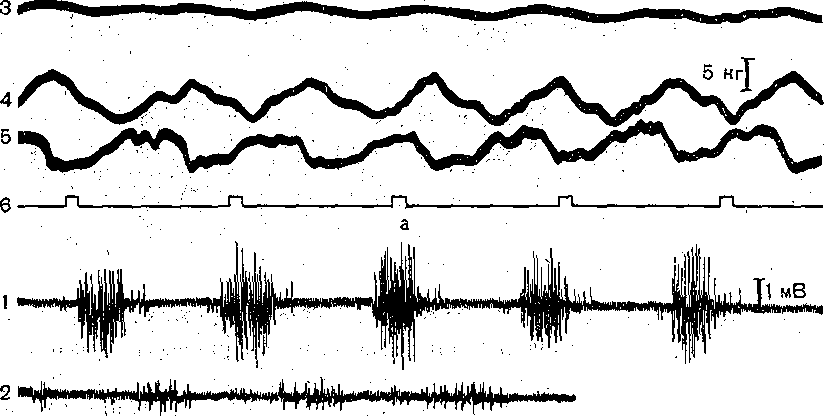 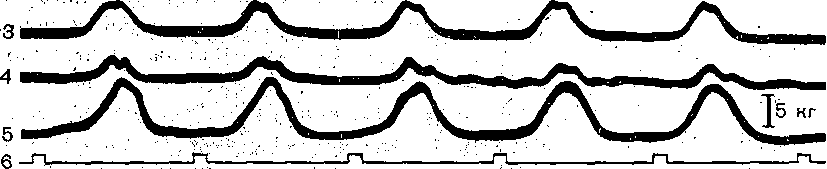 Рис.- 100. Регистрация электромирграмм и. механограмм при опиловке. а — до обучения; б — после двухнедельного обучения, 1—электромиограмма трехглавой мышцы плеча; 2 — электромиограмма двуглавой м'ышцы плеча; 3, 4, 5 — регистрация усилий, прилагаемых к напильнику, вниз, в сторону и вперед; 0 - отметка времени 1 с. изометрическому, а длительность сокращения значительна. В «позной» деятельности мышц участвуют преимущественно низкоиороговые, медленные, устойчивые к утомлению двигательные единицы. Поза и движение не существуют самостоятельно, поскольку движение всегда происходит в условиях некоторой позы, а смена поз осуществляется путем Движения. Одна из основных задач «нозной» деятельности мышц — удержание в нужном положении суставов и звеньев тела в поле силы тяжести (удержание головы-от свисания, голеностопных суставов от тыльного сгибания при.стоянии и др.). Кроме того, «позная» активность может быть направлена на фиксацию суставов, не принимающих участия в данном движении. В трудовой деятельности удержание позы бывает связано с преодолением внешних сил. Типичный пример позы — стояние человека, Сохранение равновесия при стоянии возможно в том случае, если проекция центра тяжести тела находится в пределах площади, занимаемой на плоскости опоры стопами (рис. 101). При стоянии вертикаль, иду щая через общий центр тяжести тела, проходит несколько впереди оси голеностопных и коленных суставов и несколько позади оси тазобедренных суставов. Следовательно, на эти суставы действуют моменты сил тяжести вышерасположенных звеньев тела, а это делает необходимым для удержания позы стояния напряжение многих мышц туловища и ног. Активность этих мышц невелика. Некоторые из них, например прямая мышца живота и двуглавая мышца бедра, поддерживают постоянную активность, другие, например мышцы голени, активируются периодически. Последнее связано с небольшими колебаниями центра, тяжести тела, постоянно происходящими при стоянии как в Сагиттальной, так и во фронтальной плоскости. Эти колебания могут быть зарегистрированы специальным прибором — стабйлографом, работающим с помощью тензодатчиков (рис. 102) . Мышцы гОлени противодействуют отклонениям тела, возвращая его в вертикальное положение (рис. 103). Таким образом, поддержание позы — это активный процесс, осуществляющийся, как и движение, с участием обратных связей от рецепторов. Среди последних в поддержании позы важную роль играет вестибулярный аппарат. Поза стояния у человека энергетически относительно экономна, так как моменты силы тяжести невелики вследствие близости проекции центра тяжести тела от осей основных суставов ног. Менее экономно стояние у многих животных, например, у кошки и собаки, которые стоят на полусогнутых конечностях. Поддержание постоянной позы —только частный случай «позной» активности мышц. Обычно в процессе двигательной деятельности происходит смена поз, так как движение связано с изменением положения центра тяжести тела и его звеньев. Сохранение устойчивости при движениях обеспечивается тем, что перераспределение «цозной» активности мышц обычно предшествует движению.' 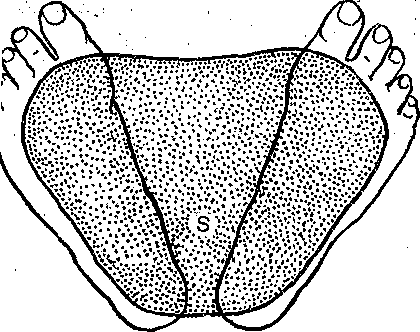 Рис. 101. Площадь опоры вольном стоянии. S |