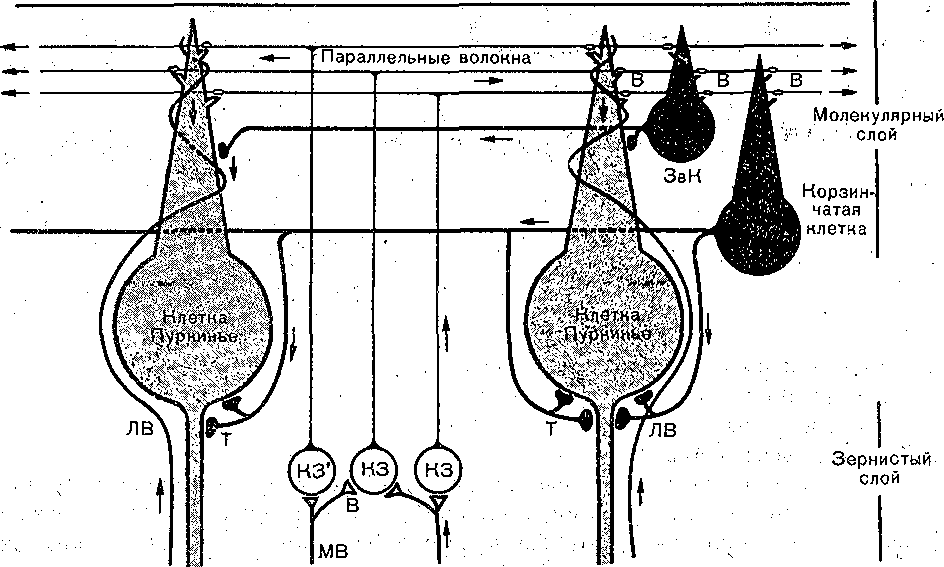
Если в кору мозжечка входит два типа афферентных волокон: лазающие й мшистые, то покидает ее всего лишь один тип эфферентных вОлокон, являющихся аксонами грушевидных нейронов (нейроны Пуркинье). Таким образом, грушевидные; клетки образуют единственный выход всей сложно организованной нейронной системы, составляющей кору мозжечка.'Другим отличительным свойством грушевидных клеток является то, что все они являются тормозными нейронами, т. е. образуют тормозящие синапсы со всеми клетками, с которыми они контактируют. Тормозные, постсинаптические потенциалы при активации грушевидных клеток возникают в нейронах собственных ядер мозжечка и в нейронах вестибулярных ядер. -
' В мозжечок поступает информация из
: различных сенсорных систем. Афферентные сигналы достигают мозжечка по различным путям, которые можно подразделить на следующие группы:
восходящие от спинного мозга (по
спинно-мозжечковым трактам) , от вестибулярных рецепторов, от
нижней оливы и от
ретикулярной формации заднего мозга.Волокна дорсального и вентрального спинно-мозжечковых трактов доставляют в мозжечок информацию о состоянии мышечного аппарата.
Спинно-ретикуломозжечковый тракт имеет переключение в латеральном ретикулярном ядре продолговатого-мозга. По этому пути в мозжечок поступает информация от кожи и более глубоких тканей. . : -;,.,
Важный афферентный'вход
кора мозжечка получает из нижней оливы, где происходит переключение импульсов, поступающих по нескольким путям, берущим начало, как-в спинном мозге, так и в структурах головного мозга. Значительную роль при этом играют сигналы, поступающие в его кору из больших, полушарий по мшистым и лазающим волокнам.
Наконец, в кору флоккулонодулярной доли приходят первичные и вторичные вестибулярные афферентные влияния. - .
Из красного ядра к мозжечку подходят коллатерали руброспинальных аксонов. Нейроны промежуточного; ядра мозжечка посылают волокна к клеткам красного ядра. Синапсы, образуемые этими волокнами на руброспинальных нейронах* являются возбуждающими и характеризуются высокой эффективностью. Поэтому раздражение промежуточного ядра мозжечка вызывает в спинальных мотонейронах ответы, сходные с теми, которые возникают при стимуляции красного ядра (рис. 91).
Нейроны других мозжечковых ядер образуют возбуждающие синапсы на ретикуло- спинальных нейронах продолговатого мозга и моста. Итак,
вся информация, приходящая в мозжечок, передается грушевидным клеткам или клеткам Пуркинье, а те в свою очередь оказывают тормозящее влияние на ядра мозжечка (а через них тормозят активность ретикуло- и руброспинальных нейронов) и на нейроны преддверного латерального ядра (ядро Дейтерса), дающие начало вестибулоспинальному тракту. Таким образом,
мозоюечок может эффективно контролировать значительную часть команд, поступающих в спинной мозг по основным нисходящим трактам. Действительно, после удаления мозжечка ритмика вестибуло-, ретикуло- и руброспинальных нейронов, которая в норме изменяется в соответствии с выполнением определенной части двигательного акта, перестает зависеть от двигательных циклов.
Функции мозжечкаНесмотря на исключительную упорядоченность нейронной организации мозжечка, благодаря которой его сравнивают со своего рода нейронной машиной, его функциональная роль раскрыта, еще не полностью.
Клинические проявления, развивающиеся при поражении мозжечка, а также эффек' ты, наблюдаемые при его раздражении или экстирпации, свидетельствуют о важной роли мозжечка в осуществлении
статических, статокинетических рефлексов и других
процессов управления двигательной активностью, автоматически регулирующих работу двигательного аппарата.
Экспериментальные и клинические наблюдения показывают, что при поражениях мозжечка развиваются разнообразные нарушения двигательной активности и мышечного тонуса, а также вегетативные расстройства. Основные проявления расстройств двигательной системы включают
нарушения равновесия и
мышечного тонуса; тремор, атаксию и
асинергию движений.Полное удаление мозжечка или егО'передней доли у животных приводит к повышению тонуса мышц-разгибателей, в то время как раздражение передней доли — к снижению этого тонуса (торможение децеребрационной ригидности).
Через несколько суток после удаления мозжечка тонус разгибателей ослабляется, сменяясь
гипотонией, лежащей в основе двигательных нарушений. В этот период животные с удаленным мозжечком не могут не только ходить, но и стоять и в то же время способны хорошо плавать.
Многие клинические проявления, свидетельствующие о мозжечковой недостаточности, связаны с мышечной
атонией и
неспособностью поддерживать позу. После исследования коленного рефлекса или смещения пассивно висящей конечности нога не возвращается в исходное положение, а раскачивается подобно маятнику.
Одно из наиболее характерных проявлений мозжечковой недостаточности — возникновение тремора. Тремор покоя характеризуется небольшой амплитудой, колебания протекают синхронно в разных сегментах тела. Для мозжечковых повреждений характерна также атаксия: нарушение величины, скорости и направления движений, что приводит к утрате плавности и стабильности двигательных реакций. Целенаправленные движения, например попытка взять предмет, выполняются порывисто, рывками, промахами мимо цели. Атаксический тремор наблюдается'при выполнении произвольных движений, будучи
наиболее выражен в начале и в конце движения, а также при перемене его направления.
Асинергия проявляется также нарушением взаимодействия между двигательными- центрами различных мышц. Так, у больных при выполнении движений не происходит одновременного сокращения мышц, компенсирующих смещение центра тяжести. Делая шаг, больной выносит ногу, не сгибая, впереди туловища, что приводит к падению. В результате асинергии сложные движения как бы распадаются на ряд выполняемых последовательно более .простых движений. ,
Разновидностью асинергии можно считать характерный для поражения мозжечка симптом —
адиадохокинез — нарушение правильного чередования противоположных движений, например сгибания и разгибания пальцев.
Итак, расстройство равновесия и тонуса скелетной мускулатуры приводит к характерным нарушениям в осуществлении произвольных движений. Особенно сильно затрудняется выполнение задач, связанных с необходимостью точно коснуться какого-либо .предмета. Нарушение двигательной координации —
астазия — характеризуемся появлением качательных и дрожательных движений. Нарушение локомоции — атаксия — проявляется расстройством походки, которая становится неровной, зигзагообразной.
Наступает неадекватное перераспределение мышечного тонуса — дистония. Локальные повреждения небольших участков коры передней доли мозжечка позволили выявить локализацию в ней представительства различных участков скелетной мускулатуры. .
Нарушение двигательной координации при поражениях мозжечка объясняется тесными связями его с основными структурами ствола мозга (дающими начало трактам, передающим импульсы в спинной мозг), а также с таламусом и сенсомоторной.областью коры больших полушарий.
;Одна из главных проекций мозжечка направлена к нейронам ретикулярной формации ствола, особенно к тем из ретикулоспинальных клеток, аксоны которых отличаются наиболее высокой скоростью проведения возбуждёния. Учитывая, что последние устанавливают контакты с а- и у-мотонейронами, можно заключить, что через ретикулоспиналь- ный путь обеспечивается срочная передача мозжечковых команд непосредственно к этим ключевым элементам спинального управления движениями.
Ретикулоспинальные нейроны находятся под мозжечковым контролем, после устранения которого участие их в центральной регуляции движений резко нарушается. .
Связи между мозжечком и преддверным латеральным ядром (ядро Дейтерса), дающим начало вестибулоспинальному тракту, настолько тесны и характерны, что это вестибулярное ядро функционально можно рассматривать как ядро мозжечка, вынесенное в продолговатый мозг. Значительная часть мозжечкового возбуждающего и тормозящего контроля спинальных центров обеспечивается с помощью вестибулоспинальных нейронов.
Команды из промежуточной коры и промежуточного ядра мозжечка передаются к спинному мозгу через нейроны красного ядра. '
! ; Таким образом, нейронная организация мозжечка обеспечивает поступление в его кору разнообразной афферентной информации, в том числе от различных компонентов двигательного аппарата, сложную обработку этой информации в нейронах и синапсах мозжечка и. эффективную передачу корригирующих влияний к нейронам стволовых и спинальных центров моторного контроля.
Как было показано Л. А. Орбели, мозжечок играет также важную роль в регуляции вегетативных функций за счет многочисленных синаптических связей с ретикулярной формацией ствола мозга!
ПРОМЕЖУТОЧНЫЙ МОЗГПромежуточный мозг образует стеной III желудочка. В процессе эмбриогенеза он формируется вместе с большими полушариями из переднего мозгового пузыря.
Главными
структурами промежуточного мозга являются шламу с, или зрительный бугор, и
гипоталамус, или подбугровая область. Ядра таламуса расположены главным образом в Области боковой стенки III желудочка; ядра гипоталамуса образуют его нижнюю и нижнебоковую стенки. Верхняя часть III желудочка образована сводом и эпифизом (эпиталамус).
vВ глубине мозговой ткани промежуточного мозга расположены ядра
наружных и
внутренних коленчатых тел. Наружная граница промежуточного мозга проходит лате- ральнее коленчатых тел и образована белым веществом внутренней капсулы, отделяющей промежуточный мозг от подкорковых ядер конечного мозга.
ТАЛАМУС
 Скачать 7.39 Mb.
Скачать 7.39 Mb.