Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
 Скачать 7.39 Mb. Скачать 7.39 Mb.
|

|
Нейронная организация. Нервные клетки таламуса, группируясь, образуют большое количество ядер: всего различают до 40 таких образований. Топографически все они могут быть подразделены на несколько основных групп: передние, интраламинарные, срединные и задние. В каждой'из этих основных групп различают более мелкие ядра, отличающиеся друг от друга как нейронной организацией, так и особенностями афферентных и эфферентных проекций. С функциональной точки зрения принято различать неспецифические и специфические ядрд таламуса. Нейроны неспецифических ядер посылают аксоны диффузно ко. всей новой, коре, в то время как нейроны специфических ядер образуют связи только с клетками определенных корковых полей (рис. 92). Неспецифические ядра являются по своему происхождению более древними и включают срединные и интраламинарные ядра, а также медиальную часть переднего вентрального ядра. Нейроны неспецифических ядер сначала передают сигналы в подкорковые структуры, от которых импул'ьсация поступает параллельно в разные отделы коры. Неспецифические ядра являются продолжением ретикулярной формации среднего мозга, представляя Собой ретикулярную формацию таламус а. ,На нейронах специфических ядер заканчиваются волокна различных восходящих трактов. Аксоны этих нейронов образуют прямые моносинаптические связи с нейронами сенсорной и ассоциативной коры. К клеткам ядер латеральной группы таламуса, включающих.заднее вентральное ядро, поступают импульсы от кожных рецепторов', двигательного аппарата, а также мозжечково-таламического пути. Другая часть специфических ядер таламуса входит в состав задней группы и образует медиальное и латеральное коленчатые тела. На нейронах латерального коленчатого тела заканчиваются афферентные пути зрительной системы. Нейроны медиального коленчатого тела воспринимают сигналы от нейронов слуховых ядер продолговатого мозга и задних бугров четверохолмия. Нейроны специфического комплекса ядер посылают по направлению к коре аксоны, почти не имеющие коллатералей. В отличие от него нейроны неспецифической системы посылают аксоны, дающие множество коллатералей. Вместе с тем и волокна, приходящие из коры к нейронам специфических ядер, характеризуются топографической л окал и.-' зованностью своих окончаний в противоположность широко разветвленной/системе диф-. фузно оканчивающихся волокон в неспецифических ядрах. 1 Функции таламуса Все сенсорные сигналы; за исключением возникающих в обонятельном тракте; достигают коры больших полушарий только;через таламокортикальные проекции. Таламус представляет собой своего рода ворота, через которые в кору поступает и достигает сознания основная информация об окружающем нас мире и о состоянии нашего тела. Тот факт, что афферентные сигналы на пути к коре мозга переключаются на нейронах'таламуса, ймеег важное значение. Тормозные влияния, приходящие в таламус из коры, других образований и соседних таламических ядер, позволяют обеспечить лучшую передачу в кору мозга наиболее важнойинформации. Торможение подавляет слабые возбуждающие влияния, благодаря чему выделяется наиболее важная информация, при1 ходящая в таламус от различных рецепторов. Через неспецифические ядра таламуса в кору мозга поступают восходящие активирующие влияния от ретикулярной формации мозгового ствола. Система неспецифических ядер таламуса осуществляет контроль ритмической активности коры больших полушарий и выполняет функции внутриталамической интегрирующей системы. Электрическое раздражение неспецифических ядер таламуса вызывает в коре больших полушарий периодические колебания потенциалов, синхронные с ритмом активности таламических структур. Реакция в коре возникает с большим скрытым периодом и значительно усиливается при повторении. Таким образом, нейроны коры больших полушарий вовлекаются в процесс активности как бы постепенно. Такая реакция вовлечения коры мозга отличается от специфических ее ответов своей' гёнерализованноетью, охватом обширных областей коры. Активацию нейронов неепецифических ядер таламуса особенно эффективно вызывают болевые сигналы (таламус является высшим центром болевой чувствительности). Импульсы, идущие по путям болевой чувствительности, формируются при раздражении различных областей тела и внутренних органов. Скрытые периоды ответов в таламусе отличаются большой длительностью и вариабельностью. Наблюдения, проведенные на людях во время хирургических операций, показывают, что повреждения неспецифических ядер таламуса приводят к нарушениям сознания. Это Рис. 93. Схематическое изображение-основных ядер гипоталамуса. I— передняя комиссура; 2 преоптиче- скоё ядро; 3 — супраоптическое ядро; 4 — ножка гипофиза; 5 — вентромедиальное гипоталамическое ядро; • 6 — дорсомеди- альное гипоталамическое ядро; 7 — медиальное ядро мамиллярного тела; 8 — ма- миллярно-таламически й пучок; 9 — заднее гипоталамическое ядро; 10 — паравентри- кулярное ядро. свидетельствует о том, что импульсация, поступающая по неспецифической восходящей системе таламуса, поддерживает уровень-возбудимости корковых нейронов, необходимый для; сохранения сознания, ; Кроме специфических влияний на кору, ряд таламических ядер, в особенности ядра дорсальной группы, оказывает регулирующее воздействие на- подкорковые, структуры. Вероятно, через эти ядра осуществляется, замыкание ;путей некоторых рефлексов, осуществляющихся без участия коры больших полушарий мозга. Таким образом, таламус может играть важную роль в качестве надсегментарного центра рефлекторной деятельности. ■ ■■■••• I,. У таламйческих животных (у которых удалены-мозговые структуры, расположенные выше таламуса) сохраняются локомоция и сложные двигательные рефлексы типа глотания, жевания, сосания.. После перерезки мозга- ниже таламуса эти функции нарушаются ГИПОТАЛАМУС Нейронная организация. Гипоталамус образован группой небольших ядер, расположенных у основания мозга, вблизи гипофиза. Клеточные ядра, образующие гипоталамус представляют собой высшие подкорковые центры вегетативной нервной системы и все> жизненно важных функций организма. В эволюционном плане гипоталамус являете* очень древним образованием. Он хорошо развит уже у наиболее примитивных представителей позвоночных. Организация гипоталамуса сохраняет сходные черты на разных ступенях эволюции, что обусловлено известным постоянством его функций; —I—;—;————:— Зрительная V ! : Слуховая i'1 Опматгу у Рис. 92. Схематическое изображение специфических и неспецифических та л а мо ко рти ка л ь - ных проекций. НП — неспецифические таламические пути; СП — Специфические таламические пути. Скопление нейронных образований, образующих гипоталамус, может быть подразделено на преоптическую, переднюю, среднюю, наружную и заднюю группы ядер. В пре оптическую область входят пёривентрикулярное, медиальное и латеральное преоптиче■ с кие ядра. В группу ядер переднего гипоталамуса относят супраоптическое, супрахиазма■ тическое и паравентрикулярное ядра. Средний гипоталамус составляет вентромедиаль- ное и дорсомедиальное ядра. К. наружной группе ядер относятся латеральное гипота- ламическое ядро и ядро серого бугра. Наконец, в заднем гипоталамусе различают заднее гипоталамическоеперифорникальное, премамиллярное, медиальное мамиллярное, суп- рамамиллярное и латеральное мамиллярное ядра. Схема главных- ядер гипоталамуса представлена на рис. 93. Организация гипоталамуса характеризуется обширными и очень сложными афферентными и эфферентными связями. - Афферентные сигналы в. гипоталамус поступают из коры больших полушарий, из таламических структур, ядер базальных ганглиев. Одним из основных эфферентных путей является медильный мозговой пучок, или паравентрикулярная система* и мамилло- тегментальный тракт. Волокна этих путей идут в каудальном направлении по стенкам водопровода мозга или сильвиева, водопровода, дают Многочисленные ответвления ж структурам среднего мозга. Аксоны клеток гипоталамических ядер образуют также большое количество коротких эфферентных путей, идущих в таламическую и субталамическую области и в другие подкорковые образования. ■ Ядра переднего гипоталамуса — супраоптическое и паравентрикулярное, кроме того, связаны с гипофизом особой системой волокон, которые служат не только^для проведения электрических сигналов, но и для транспорта продуктов нейросекреции, которые вырабатываются нейронами, этих ядер. Функции гипоталамуса Результаты, полученные с помощью избирательного раздражения или разрушения определенных ядер, показали, что латеральная и дорсальная группы ядер повышают тонус симпатической нервной системы. Раздражения области средних ядер (в частности, серого бугра) вызывают снижение тонуса симпатической нервной системы. Существуют экспериментальные данные о наличии в гипоталамусе центра сна и центра пробуждения. Гипоталамус играет важную роль в терморегуляции: Раздражение задних ядер при- юдит к гипертермии в результате повышения теплопродукции при интенсификации обменных процессов, а также вследствие дрожи скелетной мускулатуры. В области средних и боковых ядер имеются группы нейронов, рассматриваемых как центры насыщения и голода. Стимулом для изменения их деятельности являются отклонения в химическом составе притекающей крови. При голодании в крови, происходит снижение содержания аминокислот, мирных сислот, глюкозы и других веществ. Это приводит к активации определенных гипотала- лических нейронов и развитию сложных поведенческих реакций Организма, направленна на утоление чувства голода.' Приспособительные поведенческие реакции развиваются при недостатке в организме воды, что приводит к появлению чувства- жажды вследствие активации гипоталами- [еских зон, расположенных дорсолатерально. от супраоптического ядра. В результате >езко усиливается потребление воды (полидипсия). Наоборот, разрушение указанных •ипоталамических центров приводит к отказу от воды (адипсия). В гипоталамусе расположены центры,'связанные с регуляцией полового поведения. )пыты с вживлением электродов ^ эти центры (задний гипоталамус) показали, что при федоставлении животному возможности самораздражения (путем нажатия педали, жлючающей ток, проходящий через вживленные электроды) оно может проводить само- >аздражение с высокой'частотой в течение длительного времени. Поэтому эти центры бы- ш названы центрами удовольствия. Установлено, что они являются компонентом нейрон- юй системы, принимающей участие в,регуляции эмоциональной сферы полового поведения. Гипоталамус принимает участие в процессе чередования сна и бодрствования. Супраоптическое ядро и супраоптико-гипофизарный тракт связаны с задней долей 'ипофиза, выделяющей в кровь ряд гормонов. Установлено, что по крайней мере часть этих гормонов (многие из которых являются полипептидами) может выделяться окон чаниями нейронов других отделов нервной системы в качестве нейромедиаторов или ней- ромодуляторов. Задняя доля гипофиза, являющаяся по происхождению производным нервной системы (нейрогипофиз)*, специализировалась на депонировании и выделение указанных веществ в кровяное русло. Эти вещества продуцируются клетками супраопти- чес кого ядра и передаются в кровь по их аксонам в результате генерации потенциал? действия в нейрогипофиз подобно тому как потенциалы действия, приходящие в аксо нальные окончания обычных нервных клеток, вызывают процесс высвобождения ме диатора. ■ Основными гормонами, выделяемыми задней долей гипофиза, являются антидиуретический, гормон, регулирующий водный метаболизм, а также гормоны, регулирующие деятельность матки, функцию молочных желез. . По-иному осуществляется связь гипоталамуса с передней долей гипофиза (аденоги- пофиз),. вырабатывающего такие гормоны белковой природы, как адренокортикотроп- ный, фолликулостимулирующий и лютеинизирующий, тиреотропный, гормон роста, v средней долей гипофиза (меланофорный гормон). Регуляция гипоталамусом этой част* гипофиза осуществляется через кровь --нейрогуморальным путем. Важнейшие функ ции гипоталамуса приведены в табл. 6 и 7, Таблица 6 Регуляция выделения гормонов и Терморегуляция Гормоны Источники сигналов, вызывающих выделение гормонов Области гипоталамуса, ответственные за выработку гормонов 1. Катехоламины.
. лютеинизирующий гормон (ЛГ)
Регуляция Эмоциональные . воздействия, главным образом со стороны лнмбической системы ' Осморецепторы Механорецепторы гениталий матки, молочных желез Терморецепторы и другие рецепторы Эмоциональные воздействия, главным образом со стороны лимбической системы, передняя доля гипофиза, рецепторы гипоталамуса, регулирующие, уровень кортикостероидов .в крови Клетки' гипоталамуса, ответственные за регуляцию уровня эстрогенов крови Тактильные рецепторы грудной железы Источники сигналов неизвестны Дорсомедиальные и задний гипота- 'ламус Супраоптические ядра Паравентрикулярные ядра Передний гипоталамус и передне- срединная эминенция гипоталамуса Средний участок, срединная эми ненцйя гипоталамуса Срединно-задняя эминенция гипо таламуса Срединно-задняя эминенция гипоталамуса (тормозная секреция) Передне-срединная .эминенция гипоталамуса Терморегуляция Холодовые. рецепторы кожи, клетки гипоталамуса, воспринимающие изменения температуры крови Передний гипоталамус, ответственный за повышение температурь тела, задний гипоталамус, ответ ственный за снижение температуры тела Таблица 7 Регуляция мотивационного поведения и защитные реакций
ПЕРЕДНИЙ МОЗГ , ПОДКОРКОВЫЕ ЯДРА . В состав самого рострального: отдела ЦНС. — переднего мозга входят базальные '■англии и кора больших полушарий мозга. Базальные ганглии являются структурами ядерного типа, расположены внутри боль- них полушарий, между лобными долями и промежуточным мозгом. К ним относятся хвостатое ядро й "скорлупа, объединяемые под общим названием «полосатое тело», в связи : тем, что скопления нервных клеток, образующих серое вещество, чередуются с прослойками белого вещества. Вместе с бледным шаром (паллидум) они,образуют стриопалли- )арную систему подкорковых ядер. / . ; Ядра стриопаллидарной системы заметно прогрессируют в процессе эволюционного • азвития ЦНС позвоночных, достигая особенно высокого совершенства у рептилий и гтиц, у которых они становятся основным образованием переднего мозга. У млекопитаю- цих морфофункциональный прогресс переднего мозга: связан прежде всего с развитием :оры больших полушарий. Тем не менее стриопаллидарная система у млекопитающих >стается главный ядерным аппаратом переднего,моЗга, неотъемлемым компонентом провесов интеграции двигательного поведения и ряда других сложных функций. Нейронная организация. Подкорковые ядра, входящие в систему базальных ганг- [иев, отличаются чрезвычайно разнообразным клеточным составом. В бледном шаре имеется крупные нейроны, дающие начало быстропроводящим аксонам^которые.заканчивается на нервных клетках ядер промежуточного и среднего мозга. Так,-окончания этих юлокон обнаруживаются в области красного ядра и среди нейронов черной субстанции. Сроме того, в бледном шаре имеется значительное число мелких нейронов, выполняющих, ю-видимому, функции промежуточных элементов. Хвостатое ядро и скорлупа имеют сходную нейронную организацию. Они содержат лавным образом мелкие нейроны с короткими дендритами и тонкими аксонами. Кроме «елких клеток имеется небольшое число (5 % от общего состава) относительно круп- 1ых нейронов. Как крупные, так и мелкие нейроны дают начало нервным волокнам, натравляющимся в. первую очередь к клеткам бледного' шара и черной субстанции ;рис. 94). Нейроны ядер стриопаллидарной системы получают сигналы из коры больших полушарий, таламуса, ядер мозгового ствола, обонятельной луковицы. Схематически они по- ;азаны на рис. 95. Кортико-стриальные волокна берут начало практически из всей новой :оры и распределяются во всех зонах стриатума. Наибольшее число волокон берет нача- ю. от клеток сенсомоторной зоны коры, наименьшее — от клеток зрительной зоны коры. 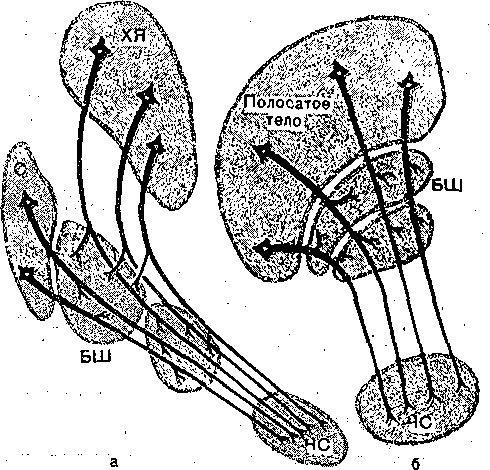 Рис. 94. Схематическое изображение топографической организации проекций нейронов полосатого тела (С) к бледному шару черной субстанции (ЧС). а дорсоаентральная а медйовентральная орга-*' ' низация; б — вентродореальная и медиолатераль; ная организация. ХЯ —хвостатое ядро. Окончания кортикальных волокон образуют синапсы преимущественно на дендритных отростках нейронов полосатого тела. Вероятным медиатором в этих синапсах является глутамат. Кортикальные волокна возбуждают нейроны стриатума. Проекции из таламуса берут начало в интраламинарных таламических ядрах. Таламостриарные проекции устанавливают синаптические контакты с шипиками или основаниями дендритов нейронов скорлупы и хвостатого тела. Это возбуждающие, синапсы; ..■.■■■;:'■ ^ ■..■.■;■..:■'■■;.. ■ . Волокна, поступающие из черной субстанции, оказывают, тормозящее влияние, на активность нейронов. - , .. ; Нейроны хвостатого ядра и скор пулы в свою очередь, дают начало большому числ> очень тонких, слабомиелинизированных волокон, которые уже в пределах этих ядер собираются в тонкие пучки и идут к бледному шару, проходя через толщу всей этой структуры. Направляясь более каудально, волокна из полосатого тела достигают черной суб станции. Значительная часть стриопаллидарных волокон представляет собой коллате ральные ответвления пути к черной субстанции. Функции подкорковых ядер . Представляя собой составную: часть экстрапирамидной системы, ядра стриопалли дарного комплекса принимают участие в координации двигательной активности. Элею рическое раздражение хвостатого ядра вызывает стереотипные движения головы, дро жательные движения-передних конечностей. Обращает на себя внимание сходство эф фектов, вызываемых электрическим раздражением ядер стриопаллидарной системы и ап Пликацией дофамина. Аппликация дофамина в область полосатого ядра вызывает те же двигательные реакции, что, и,электрическое раздражение. Вещества, .тормозящие синтез дофамина или блокаду дофаминовых рецепторов, устраняют двиГател.ьньк эффекты в ответ на электрическое раздражение хвостатого ядра. . Лобная кора 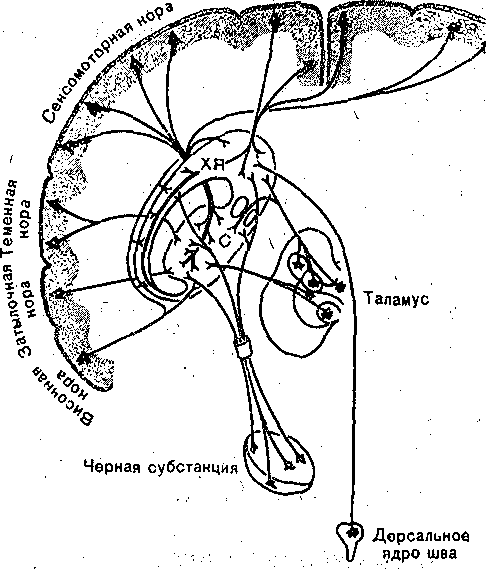 Рис. 95. Схематическое изображение организации синаптических проекций полосатого тела. Главный источник дофаминергических нейронов, аксоны которых образуют синап сы на клетках ядер Стриопаллидарн.ого комплекса, находится, в черной субстанции. По этому патологические изменения, развивающиеся в черной субстанции, ведуттс уменьшению содержания дофамина в стриатуме и нарушению его деятельности. Подобные изменения наблюдаются у больных, страдающих паркинсонизмом. Билатеральные перерезки путей, идущих из черной^субстанции в стриатум, вызывают у животных неподвижность, отказ от еды и питья, отсутствие ответов на раздражение из внешнего мира. ' : Имеются данные, что полосатое тело играет роль в процессах заяолш«аншг двигательных программ. Раздражение этой структуры может приводить к нарушению обучения, И ПаМЯТИ. ' Еще в конце прошлого века В. И; Данилевским было выдвинуто представление о то'-р1 мозной функции полосатого тела. В дальнейшем было установлено, что Стриопаллидар- ная система оказывает тормозящее влияние на различные проявления двигательной активности-и на эмоциональные компоненты двигательного поведения, в частности на агрессивные реакции. КОРА БОЛЬШИХ ПОЛУШАРИЙ ' Кора больших полушарий представляет собой филогенетически наиболее молодое образование мозга. В структурном отношении кора мозга представляет собой слой серого вещества, покрывающего весь мозг и благодаря наличию большого числа складок имеющего очень большую суммарную поверхность, : Та к, у человека поверхность коры превышает 1500 см2; Такая организация серого вещества-обеспечивает размещение громадного количества (свыше 14 млрд.) нервных клеток. Общее число образуемых ими синаптических переключений чрезвычайно велико, что обеспечивает сложнейшие процессы обработки и хранения информации. В соответствии с историей развития коры больших полушарий выделяют древнюю (архикортекс), старую (палеокортекс) и. новую кору (неокортекс). Последняя особенно быстро развивалась в процессе эволюции млекопитающих; что проявляется в значительных отличиях ме^кду новой корой приматов и более примитивных видов животных. К древней коре относят обонятельные луковицы, в которые поступают афферентные волокна от обонятельного эпителия слизистой полости носа, обонятельные тракты, расположенные на нижней поверхности лобной доли, обонятельные бугорки, в которых расположены вторичные обонятельные центры. Старая кора включает поясную извилину, извилину гиппокампа и. миндалину. Все остальные области относятся к новой коре. |