|
|
Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
Примеры отдельных видов кодирования. Код «начало — конец стимула» осуществляется в одиночном чувствительном элементе в основном на подкорковых уровнях анализатора и относится к временному кодированию. Его особенность состоит в том, что импульсация нейрона возникает только в, момент включения, выключения или резкого изменения интенсивности раздражителя. Так, например, на всех уровнях зрительного анализатора описаны нейроны, отвечающие на включение света («оп-нейроны»), на его выключение («off-нейроны»), либо же на включение и на выключение (on-off-нейроны). Такие реакции выделяют'наиболее информативные временные контуры событий. Меньшему числу периферических чувствительных клеток свойственно кодирование параметров раздражителя с помощью частоты импульсации.
Для мозга очень важен код, характеризующийся изменением распределения импуЛьсов внутри пачки или их последовательности. Импульсы могут быть по-разному сгруппированы в начале, середине или конце одной пачки импульсов, что придает ей разный временной рисунок. Для передачи сведений о сигнале имеют значение и изменения длительности пачки импульсов, число импульсов в пачке, степень колебаний частоты разрядов в их длительной последовательности. Важным кодовым признаком является последовательность включения отдельных нейронных каналов, по которым поступает информация о том, что раздражитель передвигается по рецепторной поверхности.
К кодам высокого уровня, функционирующим у высших позвоночных животных, как правило, в корковом отделе анализатора,.относится кодирование посредством последовательного включения параллельно работающих каналов, изменением.их числа, а также так называемое позиционное преобразование. Последний вид кодирования наиболее важен и заключается в том, что Определенный признак раздражителя вызывает возбуждение определенного нейрона или небольшой группы нейронов, расположенных в строго определенном месте того или иного нервного слоя. Например, возбуждение на корковом уровне зрительной системы определенной небольшой группы нейронов означает, что в определенной части поля зрения появилась световая полоска определенного размера и ориентации.
На высших уровнях анализаторов происходит переход от преимущественно временного кодирования признаков раздражителя (свойственного периферическим отделам) к преимущественно пространственному (в основном позиционному) коду.
Детектирование сигналов — специальный вид избирательного анализа, отдельных признаков раздражителя и их конкретного биологического Значения. Осуществляют такой анализ специализированные нейроны-детекторы, которые благодаря свойствам своих связе'й способны реагировать лишь на строго определенные параметры стимула.
Корковые зрительные детекторы реагируют лишь на одно из множества положений или наклонов светлой или темной полоски, расположенной в определенной части поля зрения. При другом положении той же полоски ответят другие нейроны. Совокупность нейронов, оценивающих разные стороны одного и того же признака (например, все возможные ориентации изображений), составляет систему детекции этого признака.
Общим в распределении детекторов является иерархический принцип, согласно которому на более низких уровнях локализуются детекторы более простых признаков, обеспечивающие простой анализ. В высших отделах анализатора, как правило, сконцентрированы детекторы более сложных признаков.
Опознание образов — конечная , и наиболее сложная операция •анализатора. Она заключается в классификации образа, отнесении его к тому или иному классу объектов, с которыми ранее встречался организм. Это происходит на основе всей предыдущей обработки афферентного сигнала, после расщепления его нейронами-детекторами на отдельные признаки и их раздельного параллельного анализа. Задача операции опознания может быть сведена к построению мозгом «модели раздражителя» и ее выделению из множества других подобных моделей. Опознание завершается принятием решения о том, с каким объектом или ситуацией встретился организм. Полагают, что для этого существуют специфические пространственно связанные наборы нейронов (нейронные ансамбли — высшие детекторы), возбуждение которых означает для мозга появление того или иного образа. Именно в результате такого опознания мы осознаем, какого человека видим перед собой, чей голос слышим, какой запах чувствуем и т. д.
Опознание происходит независимо от изменчивости сигнала. Мы надежно опознаем, например, предметы при различной их освещенности, окраске, размере, ракурсе, ориентации и положении в поле зрения. Точно так же знакомый голос опознается при разной его громкости, наличии звукового фона, а смысл речи — и при значительных изменениях её тембра и темпа. Отсюда следует, что на каких-то высших уровнях анализатора организуется независимое от этих изменений признаков отражение сигнала — сенсорный образ. Это совокупность сигналов, отображаемых в сходном пространственно-временном распределении процессов возбуждения и торможения на высшем уровне анализатора.
■ Взаимодействие нейронов анализаторов осуществляется с помощью двух основных механизмов — возбудительного и тормозного. Возбудительное взаимодействие представ-' лено главным образом между элементами последовательных нервных слоев, в то время как тормозное — в основном между нейронами одного и того же слоя.
Возбудительное взаимодействие организуется следующими способами. Во-первых, аксон каждого нейрона, приходя в вышележащий слой, делится на большее или меньшее число веточек, вступающих в синаптические контакты не с одним, а с несколькими нейронами.
Во-вторых, «дендритное дерево», т. е. входы нейрона, имеют, как правило, синаптические контакты с аксонами не одной, а нескольких клеток предыдущего слоя. Из-за этого практически все нейроны анализатора имеют проекционные поля, т. е. совокупность нейронов на следующем и более высоких уровнях анализатора, с которыми они взаимодействуют.
Совокупность рецепторов, импульсы от которых поступают на данный нейрон, называют его рецептивным полем.
Следует иметь в виду, что рецептивные и проекционные поля существуют у всех нейронов системы одновременно. Более того, они частично перекрываются. Такое «сдвинутое боковое перекрытие» связей заключается в том, что часть рецепторов, входящих в рецептивное поле данного нейрона, входит и в рецептивное поле соседней с ним клетки, а часть нейронов, входящих в проекционное поле какого-либо рецептора, может входить и в проекционное поле соседнего рецептора (рис. 209).
Такое сложное взаимодействие клеток приводит к образованию в анализаторе так называемой нервной сети. Благодаря ей происходит повышение чувствительности анализаторов к слабым сигналам. Кроме того, избыточность связей в сетевых структурах обеспечивает анализаторам высокую приспособляемость к меняющимся условиям среды.
Тормозное взаимодействие в анализатора.^ осуществляется с помощью тормозных вставочных нейронов (интернейронов). Чаще всего торможение заключается в том, что каждый возбужденный нейрон активирует тормозной интернейрон,'который.в свою очередь подавляет, затормаживает импульсацию как самого возбудившего его элемента, так и его соседей по слою. Сила этого торможения тем больше, чем сильнее возбужден элемент и чем ближе к нему соседняя клетка. За счет такого универсального механизма торможения в анализаторных системах осуществляется значительная часть операций по снижению избыточности информации, поступающей из рецепторов, а также по выделению сведений о пространственных и временных признаках раздражителей.
Торможение играет ведущую роль и во временной обработке сигналов, так как .включается, как правило, с некоторой задержкой после возбуждения и подавляет преимущественно поздние по отношению к началу раздражения ответы нейронов. В результате этого они хорошо отвечают только. на изменения раздражителя во времени, в частности на его включение и выключение.
АДАПТАЦИЯ АНАЛИЗАТОРОВ
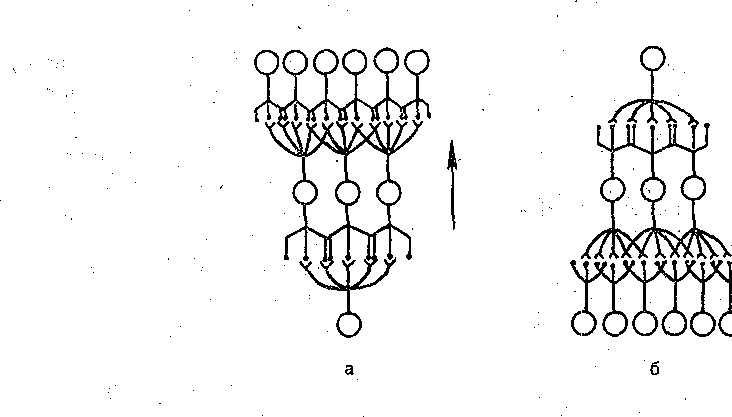
Рис. 209. Схема проекционного (а) и рецептивного (б) полей нейрона. Нервная сеть (в). Направление потока импульсов показано стрелкой. Возбужденные нейроны в последовательных слоях (1—5) выделены.
2 1
. Анализатор работает как единая система, все звенья которой взаимосвязаны и взаимно регулируют друг друга. Состояние практически всех уровней анализатора контролируется (прямо или опосредованно) ретикулярной формацией, включающей их в единую систему, интегрированную с другими отделами мозга и организмом в целом. В этой интегративной деятельности особую роль приобретает адаптация анализаторов — их общее свойство, заключающееся в приспособлении всех их звеньев к постоянной интенсивности длительно действующего раздражител я. Адаптация проявляется, во-первых,
в снижении абсолютной чувствительности анализатора, и во-вторых, повышении его дифференциальной чувствительности к стимулам, близким по силе к адаптирующему.
Субъективно адаптация проявляется в привыкании к действию постоянного раздражителя: войдя в прокуренное помещение, человек через несколько минут перестает ощущать столь резкий вначале запах табака. Точно так же мы не замечаем непрерывного давления на кожу привычной одежды или яркого света, заливающего помещение (хотя в момент, когда мы вошли в него из темной комнаты, он нас Ослепил). Повышение дифференциальной чувствительности анализатора во время адаптации заключается в том, что на фоне длительно действующего раздражителя различается больше градаций сравнительно слабых его изменений.
Адаптационные процессы начинаются на уровне рецепторов, охватывая все нейронные уровни анализатора. Адаптация заметно не изменяется только в вестибуло- и проприоцепторах. По скорости данного процесса все рецепторы делятся на быстро- и медленноадаптирующиеся. Первые после развития адаптационного процесса,,практически вообще не сообщают следующему за ними нейрону о длящемся раздражении, у вторых эта информация передается, хотя и в значительно уменьшенном виде. Когда действие постоянного раздражителя прекращается, чувствительность анализатора повышается. ТакоВа причина повышения световой чувствительности нашего глаза в темноте.
Эфферентная регуляция физиологических свойств анализатора проявляется изменением (настройкой) рецепторов и свойств нервных элементов анализатора для оптимального восприятия внешних сигналов.
Давно известен комплекс реакций (например, изменение положения тела или головы, глаз и ушных раковин по отношению к источнику звукового раздражения), оптимизирующих условия восприятия сигнала. '
В настоящее время получено много данных о преобразовании афферентного потока, идущего от рецепторов к высшим чувствительным центрам, под воздействием эфферентного контроля со стороны ЦНС. Этот контроль затрагивает элементы всех без исключения уровней анализатора, доходя до рецепторных аппаратов. Пути реализации эфферентных воздействий различны: изменение кровоснабжения рецепторов, влияние на мышечный тонус вспомогательных структур рецепторных аппаратов, на состояние самих рецепторов й нервных элементов следующих уровней. Эфферентные влияния в анализаторах чаще всего имеют тормозной характер, т. е. приводят к уменьшению их чувствительности и'ограничивают поток афферентных сигналов.
Общее число афферентных нервных волокон, приходящих к рецепторам или к элементам какого-либо нервного слоя анализатора, как правило, в десятки раз меньше числа афферентных нейронов, расположенных на том же уровне. Это определяет ёажную функциональную особенность эфферентного контроля, который имеет, не тонкий и локальный, а достаточно широкий и диффузный характер. Речь идет об общем снижении чувствительности значительной части рецепторной поверхности.
Взаимодействие анализаторов осуществляется на нескольких уровнях: спи- нальйом, ретикулярном и таламокортикальном. Особенно широка интеграция сигналов в нейронах ретикулярной формации. В коре мозга происходит интеграция сигналов высшего порядка. В результате множественных связей с нижележащими уровнями анализаторов и неспецифических систем Многие коркЬвые, Нейроны приобретают способность отвечать на сложные комбинации сигналов разной природы. В особенности это свойственно клеткам ассоциативных областей и двигательной зоны коры больших полушарий. В этой зоне пирамидные клетки служат общим конечным путем зрительных, слуховых, тактильных и других сигналов. Афферентные связи нейронов этих областей обладают высокой пластичностью, что обеспечивает их условнорефлекторные перестройки и тем самым формирование новых или видоизменение выработанных ранее навыков. Особенно важны для межсенсорного синтеза лобные доли коры больших полушарий: при их поражении у людей затрудняется формирование сложных комплексных образов."
ЧАСТНАЯ ФИЗИОЛОГИЯ АНАЛИЗАТОРОВ
ЗРИТЕЛЬНЫЙ АНАЛИЗАТОР
Зрительный анализатор (или зрительная сенсорная система) — важнейший из органов чувств человека и большинства высших позвоночных животных. Он дает более 90% информации, идущей к мозгу от эсех рецепторов (не случайна пословица «Лучше один раз увидеть, чем сто раз услышать»). Благодаря опережающему эволюционному развитию именно зрительных механизмов мозг хищных животных и приматов претерпел резкие изменения и достиг значительного совершенства. Зрительное восприятие — многозвеньевой процесс, начинающийся с проекции изображения на сетчатку глаза и возбуждения фоторецепторов и заканчивающийся принятием высшими отделами зрительного анализатора, локализованными в коре мозга, решения о наличии в поле зрения того или иного зрительного образа.
Оптическая система глаза. В связи с необходимостью наводить глаз на рассматриваемый объект, вращая его, природа создала у большинства видов животных шарообразную форму глазного яблока. На пути к светочувствительной оболочке глаза — сетчатке—лучи света проходят через несколько прозрачных поверхностей — переднюю и заднюю поверхности роговицы, хрусталика и стекловидного тела. Разная кривизна и показатели преломления этих поверхностей определяют преломление световых лучей внутри глаза.
Преломляющую силу любой оптической системы выражают в диоптриях (D). Одна диоптрия равна преломляющей силе линзы с фокусным расстоянием. 100 см. Преломляющая сила глаза человека составляет 59 D при рассматривании далеких и 70,5 D при рассматривании близких предметов.
-, т. е. изображение на сетчатке (аЬ) во столько раз меньше самого предме
та М/?)'.' во сколько dO (15 мм) меньше расстояния предмета от глаза (DO).
ь d
а
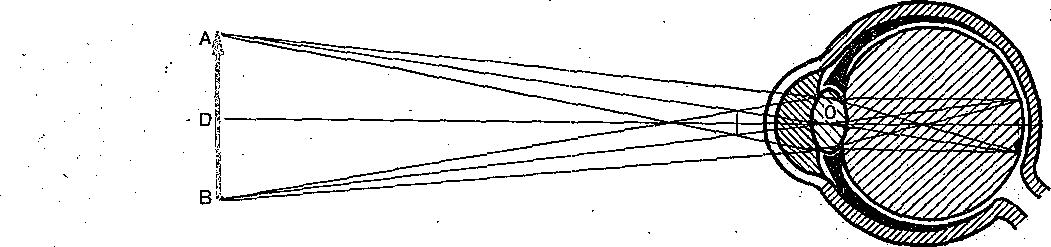
Рис. 210. Ход лучей и построение изображения в редуцированном глазу.
АВ — предмет; ab — его изображение; О — узловая точка; D — d — главная оптическая ось.
Сложность оптической системы глаза затрудняет точную оценку хода лучей внутри него .и оценку размера изображения на сетчатке. Поэтому пользуются упрощенной моделью — «редуцированным глазом», в котором все преломляющие среды имеют один и тот же показатель преломления в единую сферическую поверхность. Для того чтобы построить на сетчатке такого схематического глаза изображение, нужно провести линии от концов рассматриваемого предмета к узловой точке, лежащей в 7,15-10
3 м.(7,15 мм) кзади от роговой оболочки и на 15 мм (15- Ю
3 м) вперед от сетчатки. При этом изображение на сетчатке получается действительным, уменьшенным и обратным (рис. 210). Для построения изображения на сетчатке нужно знать величину предмета и его расстояние от роговицы глаза. Из подобия треугольника АВО и аЬО на рис. 210 ясно, что
Рис. 211. Ход лучей от близкой и далекой точки.
От далекой точки А (параллельные лучи)-изображение а получается на сетчатке при ненапряженном аккомодационном аппарате; при этом от близкой точки В изображение в образуется за сетчаткой.
Аккомодация
Для ясного видения предмета необходимо, чтобы лучи от его точек попадали на поверхность сетчатки, т. е. были здесь"сфокусированы (рис. 211). Когда человек смотрит на далекие предметы, их изображение сфокусировано на сетчатке, и они видны ясно. При этом близкие предметы видны неясно, их изображение на сетчатке расплывчато, так как лучи от них собираются за сетчаткой. Видеть одновременно одинаково ясно предметы, удаленные от глаза на разное расстояние, невозможно. В этом легко/убедиться: переводя взгляд с близкого на далекие предметы, вы перестаете его ясно видеть.
Приспособление глаза к ясному видению удаленных на разное расстояние предметов называется аккомодацией (аккомодацию глаза не следует смешивать с аккомодацией возбудимой ткани). При аккомодации происходит изменение кривизны хрусталика и, следовательно, его преломляющей способности. При рассматривании близких предметов хрусталик делается более выпуклым (рис. 212), благодаря чему лучи, расходящиеся от светящейся точки, сходятся на сетчатке.
Радиус кривизны передней поверхности хрусталика при аккомодации уменьшается с 10 до 6 мм (с Ы0
2 м до 6-10
3 м), а задней поверхности с 6 до'5,5 мм (с 6-10"3 м до 5,5• 10
3 м).
Механизм аккомодации сводится к сокращению ресничных мышц, которые изменяют выпуклость хрусталика. Хрусталик заключен в тонкую прозрачную капсулу, переходящую по краям в волокна цинновой связки, прикрепленной к ресничному телу. Эти волокна всегда натянуты и растягивают капсулу, сжимающую и уплощающую хрусталик. В ресничном теле находятся гладкомышеч- ные волокна. При их сокращении тяга
Рис. 212. Механизм аккомодации (по Гельм- гольцу).
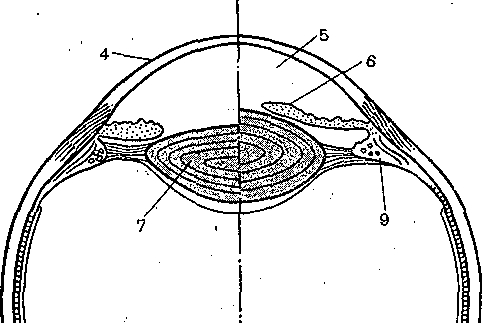
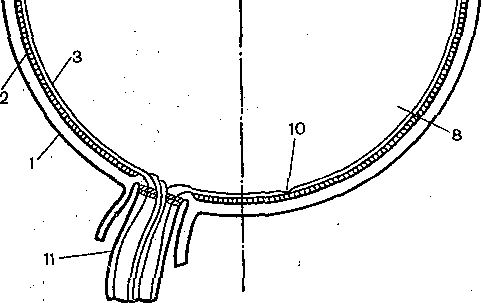
В левой половине хрусталик (7) уплощен при рассматривании далекого предмета, а справа он стал более выпуклым за счет аккомодационного усилия при рассматривании близкого предмета. I —г склера; 2 — сосудистая оболочка; 3 — сетчатка; 4 — роговица; 5 — передняя камера; 6 — радужная оболочка; 7— хрусталик; 8 — стекловидное тело; 9 — ресничная мышца, ресничные отростки и ресничные связки (цинновы); 10— центральная ямка; 11 — зрительный нерв.
цинновых связок ослабляется, а значит уменьшается давление на хрусталик, который вследствие своей эластичности принимает более выпуклую форму. Таким образом, ресничные мышцы являются аккомодационными мышцами. Они иннервируются парасимпатическими волокнами глазодвигательного нерва. Введение в глаз атропина вызывает нарушение передачи возбуждения к этой мышце, и, следовательно, ограничивает аккомодацию глаз при рассмотрении близких предметов. Наоборот, парасимпатомимети- ческие вещества — пилокарпин и эзерин — вызывают сокращение этой мышцы.
Для нормального глаза молодого человека дальняя точка ясного видения лежит в бесконечности. Далекие предметы он рассматривает без всякого напряжения аккомодации, т. е. без сокращения ресничной мышцы. Ближайщая точка ясного видения находится на расстоянии 10 см от глаза. Предметы, расположенные ближе 10 см, не могут быть ясно видны человеком с нормальным зрением даже при максимальном сокращении ресничной мышцы, т. е. при максимальном аккомодационном усилии. Сила аккомодации может быть выражена в диоптриях.
Если ближайшая точка ясного видения находится на расстоянии 10 см от глаза человека, то линза с фокусным расстоянием в 10 см, т. е. в 10 D, превратит лучи, идущие от ближайшей ясно видимой точки, в параллельные. Следовательно, при помощи линзы устранится необходимость в аккомодации. Поэтому можно заменить максимальное аккомодационное усилие человека, поместив у него перед глазом линзу, силой в 10 D, отсюда следует, что максимальная сила аккомодации равна 10 D.
|
|
|
 Скачать 7.39 Mb.
Скачать 7.39 Mb.