|
|
Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
Костная передача звуков. Кроме воздушной передачи звука через барабанную перепонку и слуховые косточки, возможна передача через кости черепа. Если поставить ножк> камертона на темя или сосцевидный отросток, звук будет слышен даже при закрытом слуховом проходе. Звучащее тело вызывает колебания костей черепа, которые вовлекают в колебание слуховой рецепторный аппарат.
Внутреннее ухо и восприятие звуков
Во внутреннем ухе, кроме преддверия и полукружных каналов (вестибулярный рецепторный аппарат), находится улитка, где расположены рецепторы, воспринимающие звуковые колебания.
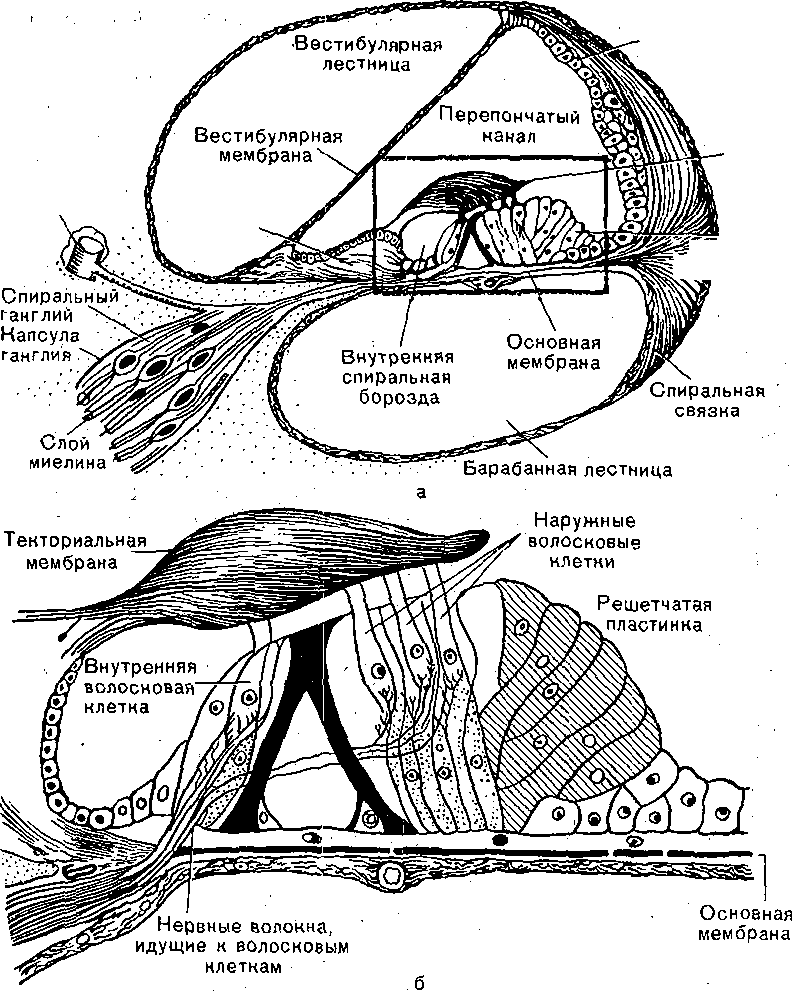
Строение улитки. Улитка представляет собой костный спиральный, постепенно расширяющийся канал, образующий у человека 2,5 витка. Диаметр костного канала у основания улитки 0,04 мм (4-10
5 м), а на вершине ее — 0,5 мм (5-10
4 м). По всей длине, почти до самого конца улитки, костный канал разделен двумя перепонками: более тонкой — вестибулярной мембраной (мембрана Рейснера) и более плотной и упругой — основной мембраной. На вершине улитки обе эти мембраны соединяются и в них имеется отверстие — геликотрема (helicotrema). Вестибулярная и основная, мембраны разделяют костный канал улитки на три узких хода: верхний, средний и нижний (рис. 221).
Верхний канал улитки, или вестибулярная лестница (scala vestibuli), берет начало от овального окна и продолжается до вершины улитки, где он через отверстие сообщается с нижним каналом улитки — барабанной лестницей (scala tympani), которая начинается в области круглого окна. Сообщающиеся через геликотрему верхний и нижний каналы представляют собой как бы единый канал, начинающийся овальным и заканчивающийся круглым окном. Верхний и нижний каналы улитки заполнены перилимфой, напоминающей по составу спинномозговую жидкость. Перилимфа каналов отделена от воздушной полости среднего уха мембранами овального и круглого окон.
Между верхним и нижним каналами, т.е. между вестибулярной и основной мембраной, проходит средний — перепончатый канал (scala media). Полость этого, канала не
сообщается с полостью других каналов улитки и заполнена'эндолимфой. Эндолимфа продуцируется специальным сосудистым образованием (stria vascularis), которое находится на наружной стенке перепончатого канала. Состав эндолимфы отличается от состава перилимфы примерно в 100 раз большим содержанием ионов калия и в 10 раз меньшим содержанием ионОв натрия. Поэтому эндолимфа заряжена положительно по отношению к перилимфе.
Внутри среднего канала улитки на основной мембране расположен звуковоспри- нимаюший аппарат — спиральный (кортиев) орган, содержащий рёцепторные волоско- вые клетки; Эти клетки трансформируют механические колебания в электрические потенциалы,, в результате чего возбуждаются волокна слухового нерва.
. Передача звуковых колебаний по каналам улитки. Звуковые колебания передаются стремечком на мембрану овального окна и вызывают колебания перилимфы в верхнем и нижнем каналах улитки. Колебания перилимфы доходят до круглого окна и приводят к смещению мембраны круглого окна наружу в полость среднего уха.
Секреторный эпителий . .
Спиральная артерия
Текториальная мембрана
Рис. 221. Поперечный разрез завитка улитки (а) с увеличенной частью спирального (кортиева) органа (б), очерченной сверху прямоугольником.
Спиральный __ орган
^^(кортиев орган)
Вестибулярная мембрана очень тонка, поэтому жидкость в верхнем и среднем кана
лах колеблется так, как будто она не разделена мембраной и оба канала являются единым общим каналом..
Упругим элементом, отделяющим этот как бы общий верхний канал от нижнего, является основная мембрана. Звуковые колебания., распространяющиеся по перилимфе и эндолимфе верхнего и среднего каналов по типу бегущей волны, приводят в движение эту мембрану и через нее могут передаваться на перилимфу нижнего канала.
Расположение и структура рецепторных клеток спирального (кортиевого) органа. На основной мембране расположены два. вида рецепторных волосковых клеток: внутренние и наружные, отделенные друг от друга кортиевыми дугами.
Внутренние волосковые клетки располагаются в один ряд; общее число их по всей длине перепончатого канала достигает 3500. Наружные волосковые клетки располагаются в 3—4 ряда; общее число их составляет 12 ООО—20 ООО. Каждая волосковая клетка имеет удлиненную форму. Один полюс клетки фиксирован на основной мембране; второй ее полюс находится в полости перепончатого канала улитки. На конце этого полюса рецепторной клетки имеются волоски: их число на каждой внутренней клетке составляет 30—40 и они очень короткие — 4—5 мкм (4—5- Ю^6 м); на каждой наружной клетке число волосков достигает 65—120, они тоньше и длиннее. Волоски рецепторных клеток омываются эндолимфой и контактируют с покровной пластинкой, или текториальной мембраной, которая по всему ходу перепончатого канала расположена над волосковыми клетками.
При действии звуков основная мембрана начинает колебаться, волоски рецепторных клеток касаются текториальной мембраны и деформируются. Это вызывает генерацию электрических потенциалов, а затем через синапсы — возбуждение волокон слухового нерва.
Электрические явления в улитке. При отведении электрических потенциалов от разных частей улитки исследователи обнаружили 5 различных электрических феноменов. Два из них — мембранный потенциал слуховой рецепторной клетки и потенциал эндолимфы — не обусловлены действием звука (они наблюдаются и при отсутствии звуковых раздражений). Три. электрических явления—.микрофонный потенциал улитки, суммационный потенциал и потенциалы слухового нерва — возникают под влиянием звуковых раздражений (рис. 222).
Мембранный потенциал слуховой рецепторной клетки регистрируется при введении в нее микроэлектрода. Так же как и в других нервных или рецепторных клетках, внутренняя поверхность мембран слуховых рецепторов заряжена отрицательно (—80 мВ). Так как волоски слуховых рецепторных клеток омываются положительно заряженной эндолимфой (-{—SO мВ), то между внутренней и наружной поверхностью их м-ембраны разность потенциалов достигает 160 мВ (80+80 мВ). Значение столь большой разности потенциалов состоит, в том, что она резко облегчает восприятие слабых звуковых колебаний. '
Потенциал эндолимфы, регистрируемый при введении.одного электрода в перепончатый канал, а другого — в область круглого окна, обусловлен деятельностью сосудистого сплетения (stria vascularis) и зависит от интенсивности окислительных процессов. При нарушениях дыхания или подавлении тканевых окислительных процессов цианидами потенциал эндолимфы падает или даже исчезает.
Если ввести в улитку электроды, соединить их с усилителем и громкоговорителем и воздействовать звуком, то громкоговоритель точно воспроизводит этот звук. Описываемое1 явление получило название микрофонного эффекта улитки, а регистрируемый электрический потенциал назван кохлеарным микрофонным потенциалом. Доказано, что он генерируется на мембране волосковой клетки в результате деформации волосков.
Частота микрофонных потенциалов соответствует частоте звуковых колебаний, а амплитуда этих потенциалов в определенных границах пропорциональна интенсивности звука, действующего на ухо.
В. ответ на сильные звуки большой частоты (высокие тона) отмечают стойкий сдвиг исходной разности потенциалов. Это явление получило название суммационного потен
циала. Различают положительный и отрицательный суммационные потенциалы. Их величины пропорциональны интенсивности звукового давления и прижатия волосков рецепторных клеток покровной мембраной.
Микрофонный и суммационный потенциал рассматривают как рецепторные потенциалы вол ос ковы х клеток. Имеются указания, что отрицательный суммационный потенциал связан с внутренними, а микрофонный и положительный суммационные потенциалы — с наружными волосковыми клетками.
И наконец, в результате возникновения в волосковых клетках при действии на них звуковых колебаний микрофонного^ суммационного потенциалов происходит импульсное возбуждение волокон слухового нерва (рис. 223). Передача возбуждения с вблоско- вой клетки на нервное волокно происходит, по-видимому, как электрическим, так.и химическим путем. . . ^
Электрическая активность путей и центров слухового анализатора
Для волокон слухового нерва даже в тишине характерна сравнительно высокая частота спонтанных разрядов (фоновой импульсации) — до 100 имп/с. При звуковом раздражении частота импульсации в волокнах нарастает в течение всего времени, пока действует звук. Степень учащения разрядов различна у разных волокон и' связана с интенсивностью воздействия. Для каждого волокна слухового нерва может быть найдена так называемая оптимальная частота звука, дающая наибольшую частоту разрядов и наиболее низкий порог реакции. Эта оптимальная частота определяется местом на основной мембране, где расположены рецепторы, связанные с данным волокном. Таким образом, частотная избирательность волокон слухового нерва отражает пространственное кодирование информации в улитке, определяемое ее конструкцией.
Рисунок возбуждения нейронов центральных отделов слухового анализатора весьма разнообразен. Наиболее часто здесь встречаются клетки, возбуждение которых длится в .течение всего времени действия стимулов, несколько снижаясь по частоте. Реже, чем в зрительном анализаторе, на низких уровнях в слуховом анализаторе встречаются нейроны, отвечающие лишь на включение и выключение звука или резкие перепады его интенсивности (on-, off-, on — о//-типа). На более высоких уровнях слухового анализатора процент таких нейронов возрастает. В слуховой зоне коры имеется много нейронов, вызванные разряды которых длятся десятки секунд после прекращения звука.
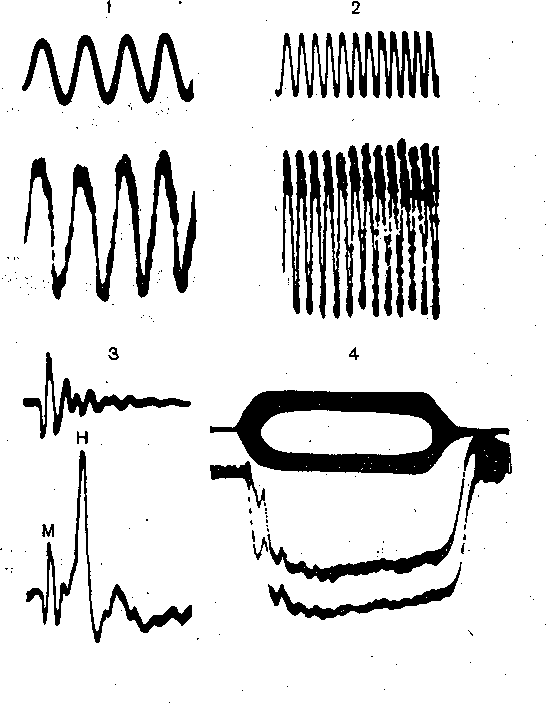
Рис. 222. Электрические явления в улитке.
Реакции, регистрируемые с круглого окна улитки (нижние кривые) в ответ на звук (верхние кривые): тон 300 Гц (1),тон 1000 Гц (2), щелчок (3), тональную посылку 21 кГц (4).
М— микрофонный потенциал; Н — нервный компонент (суммарный синхронизированный ответ волокон слухового нерва); 4 — суммационный потенциал.
На каждом из уровней слухового анализатора с помощью макроэлектродов могут быть зарегистрированы характерные пб форме вызванные 'потенциалы, отражающие синхронизированные реакции больших групп нейронов и волокон (рис. 224). -
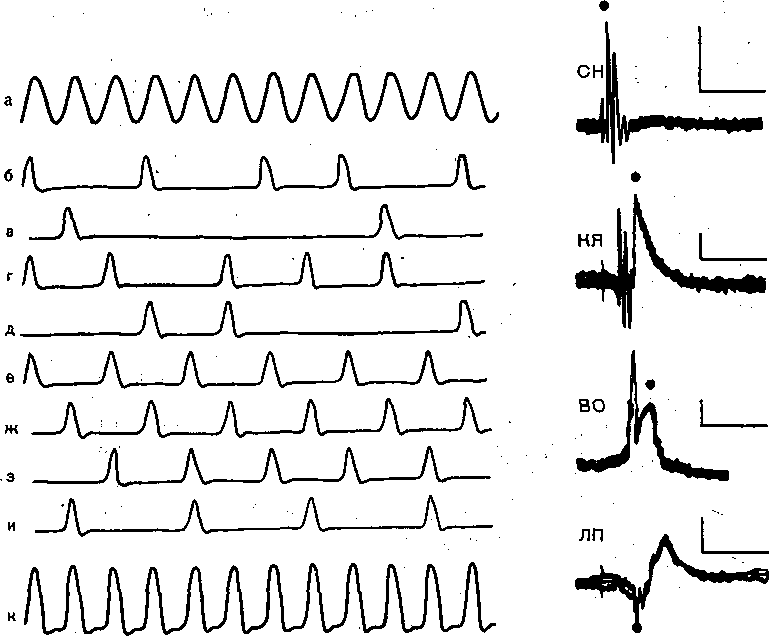
Рис. 223. Импульсация волокон слухового нерва. Схематическая иллюстрация принципа залпов (по Шварцкопфу).
а — звуковой сигнал; б — и — реакции отдельных волокон; к — суммарная реакция этих волокон, повто ряющзя частоту звука.
Рис. 224. Вызванные потенциалы разных уровней слуховой системы.
Суммарные ответы на звуковой щелчок в слуховом нерве (СН), кохлеарных ядрах (КЯ), верхней оливе (ВО) латеральной петле (ЛП), задних холмах четверохолмия (ЗХ), внутреннем коленчатом теле (ВКТ) ислухово коре (СК). Отметка времени — 5 мс слева и 20 мс справа.
Анализ частоты звуков (высоты тонов)
Звуковые колебания вовлекают в колебательный процесс перилимфу верхней и нижнего каналов улитки на всем их протяжении неодинаково. Колебательная реакции основной мембраны, передаваемая на эндолимфу, имеет характер бегущей волны, i локализация амплитудного максимума этой волйы на мембране зависит от частоть звука. Таким образом, в процесс возбуждения вовлекаются разные клетки спиральногс (кортиева) органа, примыкающие к основной мембране.
Существенное значение для восприятия звуков разной частоты локализации рецеп торных клеток спирального органа, вовлекаемых в возбуждение, доказывается, в част ности, и поведенческими экспериментами. Если повредить у собаки завиток улитю у ее основания, то исчезают ранее выработанные условные рефлексы, на высокие тона если же повреждение нанести в области вершины улитки, исчезают условные рефлексь на низкие тона; разрушение только среднего завитка улитки вызывает выпадение tohoi средней частоты диапазона. Раздельное пространственное представительство зон возбуждаемых при действии'звуков разной частоты, подтверждается электрофизиоло гическими методами (вызванные потенциалы, нейронная активность)' на всех уровня: слухового анализатора.

pLf
Для сравнительно низких частот возможен анализ высоты тона за счет реакции группы волокон слухового нерва, повторяющей частоту действия раздражителя. Этс означает, что существуют два сочетающихся механизма различения высоты тонов. Пр*
действии высоких тонов происходит лишь пространственное кодирование, основанное на неодинаковом расположении возбужденных рецепторных клеток на основной мембране. При .низких и средних тонах осуществляется и временное, .кодирование, когда информация передается по определенным группам волокон слухового нерва в виде импульсов, частота которых соответствует частоте воспринимаемых улиткой звуковых колебаний.
Основным отражением частотной настройки отдельных нейронов на всех уровнях слуховой системы является наличие у них так называемых частотно-пороговых показа-. телей. Эти показатели отражают зависимость пороговой интенсивности звука, необходимой для возбуждения клетки, от его частоты. В обе стороны по диапазону частот от оптимальной, или характеристической, частоты порог реакции нейрона резко возрастает. Таким образом, элемент оказывается «настроенным» на выделение из всей совокупности звуков лишь определенного, достаточно узкого участка частотного диапазона. Частотно- пороговые кривые разных клеток перекрывают весь частотный диапазон слышимых звуков. Форма этих кривых у многих нейронов высших слуховых центров значительно усложняется.
Анализ силы звуков (интенсивности звучания)
Сила звука кодируется .числом возбужденных нейронов и частотой их импульсации. Увеличение числа возбужденных нейронов при действий все более громких звуков связано с тем, что нейроны слухового анализатора резко отличаются друг от друга по уровню возбудимости {порогам реакций). При ;слабых раздражителях в реакцию вовлекается лишь небольшое число наиболее, чувствительных нейронов, а при усилении стимулов возбуждается все большее количество дополнительных нейронов. Кроме того, сами пороги возбуждения внутренних и наружных рецепторных клеток спирального органа неодинаковы. Возбуждение внутренних волосковых клеток возникает при большей силе звукового раздражения. Поэтому в зависимости от интенсивности звукового раздражения изменяется соотношение числа возбужденных внутренних и наружных волосковых клеток.
Звуковые ощущения
Тональность (частота) звука. Человек может воспринимать звуки с частотой колебания от 16 до 20 ООО Гц. Этот диапазон соответствует 10—11 октавам. Верхняя граница воспринимаемых звуков зависит от возраста: чем человек старше, тем она ниже;.старики часто не слышат высоких тонов (например, звука,.издаваемого сверчком). У многих животных верхняя граница слуха лежит значительно выше: у собаки, например, удается получить условные рефлексы на очень высокие, неслышимые человеком звуки. Различение частоты звука характеризуется тем минимальным различием по частоте двух звуков, которое еще улавливается человеком. При низких и средних частотах человек способен заметить различия в 1—2 Гц. Встречаются люди с абсолютным слухом: они способны точно узнавать и обозначать любой звук даже при отсутствии звука сравнения.
Слуховая чувствительность. Минимальную силу звука, слышимого человеком в половине случаев его предъявления, называют абсолютной слуховой чувствительностью. Установлено, что пороги слышимости сильно изменяются в зависимости от частоты звука.
В области частот от 1000 до 4000 Гц слух человека обладает максимальной чувствительностью. В этих пределах слышен звук, имеющий ничтожную энергию порядка 1 • 10
12 Вт/м2 (1 • Ю-9 эрг/с-см2). При звуках ниже 1000 и выше 4000 Гц чувствительность резко уменьшается: например, при 20 и при 20 000 Гц пороговая энергия звука должна быть около Ы0"3 Вт/м2 (1 эрг/с-см2) (нижняя кривая AEFGD на рис. 225).
При увеличении силы звука неизменной частоты можно дойти до7"акой силы, когда звук вызывает неприятное ощущение давления и даже боли в ухе. Звуки такой силы
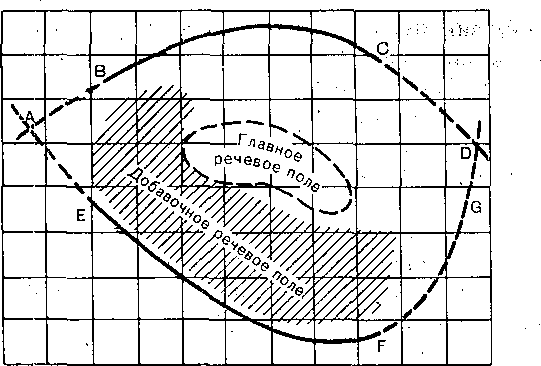
12,5 25 50 ЮО 200 400 800 1600 3200 6400 12800 2560С
Рис. 225. Область. звукового вое- _ joooo приятия человека.
„ •'•.••■ „ 1000 Зависимость пороговой интенсивности
звука (ось ординат— звуковое давле-
ние в дин/см2) от частоты тональных
звуков (ось абсцисс 'в- Гц). Линия
AEFGD абсолютные пороги, ABC0 — 10
пороги болевого ощущения от громких
звуков. '
О 1 0.01
" -г - 0.001 0.0001
дадут, очевидно, верхний предел слышимости (кривая ABGD на рис. 225) и ограничат область слухового восприятия. Внутри этой области лежат и так называемые речевые поля,, в пределах которых по частоте и интенсивности распределяются звуки речи. |
|
|
 Скачать 7.39 Mb.
Скачать 7.39 Mb.