Физиология человека. Косицкий. Литература москва Медицина 1985 Для студентов медицинских институтов
 Скачать 7.39 Mb. Скачать 7.39 Mb.
|

|
Глава 17 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ Полушария'большого мозга — их кора и ближайшие к ней подкорковые образования — являются высшим отделом центральной нервной системы (ЦНС) позвоночных животных и человека. Функции этого отдела — осуществление сложных рефлекторных реакций, составляющих основу высшей нервной деятельности (поведения). организма. Впервые представление о рефлекторном характере деятельности высших отделов головного мозга было широко и подробно развито основоположником отечественной физиологии И. М. Сеченовым в его книге «Рефлексы головного мозга». До И. М. Сеченова физиологи и неврологи не решались даже поставить вопрос о возможности объективного, чисто физиологического анализа психических процессов, которые оставались полностью отданными на откуп субъективной психологии. Идеи И. М. Сеченова получили блестящее развитие в замечательных трудах И. П. Павлова, открывшего пути объективного экспериментального исследования функций коры большого мозга, разработавшего метод условных'рефлексов и создавшего учение о высшей нервной деятельности. И. П. Павлов показал, что в то время как в нижележащих отделах ЦНС — подкорковых ядрах, мозговом стволе, спинном мозге — рефлекторные реакции осуществляются врожденными, наследственно закрепленными нервными путями, в коре большого мозга нервные связи вырабатываются заново в процессе индивидуальной жизни животных и человека в результате сочетания бесчисленных, действующих на организм и воспринимаемых корой раздражений. Открытие этого факта позволило разделить всю совокупность рефлекторных реак дий, происходящих в организме, на две основные группы — безусловные и условны* рефлексы. ОБЩАЯ ХАРАКТЕРИСТИКА И СВОЙСТВА УСЛОВНЫХ РЕФЛЕКСОВ РАЗЛИЧИЯ БЕЗУСЛОВНЫХ И УСЛОВНЫХ РЕФЛЕКСОВ Безусловные рефлексы — это врожденные, наследственно передающиеся реакцщ организма. Условные рефлексы — реакции, приобретенные организмом в процессе индивидуального развития на основе «жизненного опыта». Безусловные рефлексы являются видовыми, т.е. свойственными всем представителя!У данного вида. Условные же рефлексы индивидуальны: у одних представителей одногоvтого же вида они могут быть, а у других отсутствуют. • Безусловные рефлексы относительно постоянны, условные рефлексы непостоянны vв зависимости от определенных условий могут выработаться, закрепиться или исчезнуть; это свойство отражено в самом их названии. Осуществляются безусловные рефлексы в ответ на адекватные раздражения, приложенные к определенному рецептивному полю. Условные рефлексы могут образоваться на любые воспринимаемые организмом раздражения любого рецептивного поля. Условные рефлексы являются преимущественно функцией коры большого мозга Безусловные рефлексы могут осуществляться на уровне спинного мозга и мозгового ство ла. Они входят в состав сформированного в процессе филогенеза и наследственно передающегося фонда рефлекторных реакций. Условные рефлексы вырабатываются на базе безусловных. Для образования условного рефлекса необходимо сочетание во времени какого-либо изменения окружающей среды (или внутреннего состояния организма), воспринятого корой полушарий большогс мозга, с осуществлением того или иного безусловного рефлекса. Такой раздражитель вызывающий образование условного рефлекса, называют условным раздражителем, илк условным сигналом, в отличие от безусловного раздражителя, вызывающего безуслов ный рефлекс. КЛАССИФИКАЦИЯ БЕЗУСЛОВНЫХ И УСЛОВНЫХ РЕФЛЕКСОВ Всю совокупность безусловных "и образованных на их основе условных рефлексов по их биологическому значению принято разделять на пищевые, оборонительные, половые, статокинетические и локомоторные, ориентировочные, поддерживающие гомеостаз и некоторые другие. * ' Особое место среди безусловных рефлексов занимает ориентировочный рефлекс. Это рефлекс на новизну. Он возникает в ответ на любое достаточно быстро происходящее изменение окружающей среды и выражается внешне в настораживании, прислушивании, обнюхивании, повороте глаз и головы, а иногда и всего тела в сторону появившегося нового раздражителя и т.п. Осуществление этого рефлекса обеспечивает лучшее восприятие действующего агента и имеет важное приспособительное значение. И. П. Павлов образно назвал- ориентировочную реакцию рефлексом — «Что такое?». Реакция эта врожденная и не исчезает при полном удалении коры полушарий большого мозга у животных; ее наблюдают у детей с недоразвитыми полушариями — анэнцефалов. Отличием ориентировочного рефлекса от других безусловнорефлекторных реакций является то, что он сравнительно быстро ослабляется — угасает при повторных применениях одного и того же раздражителя. Эта особенность ориентировочного рефлекса зависит от коры большого мозга. v ' 481 Приведенная классификация рефлекторных реакций близка к классификации различных инстинктов, которые также, делят на пищевые, половые, родительские, оборони- 16 Физиология человека гельные. Это понятно в связи с тем, что инстинкты — сложные безусловные рефлексы (И. П. Павлов). Их отличительными особенностями являются цепной характер реакций (завершение одного рефлекса служит сигналом следующего рефлекса) и их зависимость эт гормональных и метаболических факторов. Так, возникновение полового и родительского инстинктов связано с циклическими изменениями функционирования половых же- чез, а пищевой инстинкт находится в зависимости от тех изменений обмена веществ, которые развиваются при отсутствии пищи. Одной из особенностей инстинктивных реакций является также то, что они характеризуются многими свойствами доминанты. КОМПОНЕНТЫ БЕЗУСЛОВНОГО И УСЛОВНОГО РЕФЛЕКСОВ Каждый безусловный рефлекс представляет собой сложную реакцию, в состав которой входит много компонентов. Условные рефлексы, как и безусловные, на базе которых )ни выработаны, также представляют собой многокомпонентную реакцию.: Среди компонентов условного рефлекса различают главные, специфические для данного вида рефлексов, и второстепенные, неспецифические компоненты. В оборонительном эефлексе главным является двигательный компонент, в пйщевом — двигательный и секреторный. ' Двигательные компоненты условных пищевых и оборонительных рефлексов отли- 1аются большим многообразием и зависят от характера безусловного раздражителя л от того, на какие рецепторы он действует. При исследовании оборонительных условных эефлексов чаще всего регистрируют сгибание конечности в ответ на безусловное электроюжное раздражение. При исследовании пищевых условных рефлексов и у людей, и у животных регистрировались такие двигательные реакции, как схватывание пищи, глотание и сосание. При сигналах, вызывающих условный оборонительный рефлекс,наблюдают условно- рефлекторное учащение и усиление сердечных сокращений. Сочетание какого-либо индифферентного раздражителя с надавливанием на глазные яблоки (что вызывает замед- иение сердечной деятельности — рефлекс Ашнера) приводит к тому, что применение только одного условного раздражителя замедляет сердечные сокращения. Возможность условнорефлекторных изменений дыхания как у животных, так и человека уже давно показана в лаборатории В. М. Бехтерева также при выработке условных эборонительных рефлексов. Условнорефлекторное увеличение легочной вентиляции и поглощения кислорода возникает в ответ на действие условного сигнала, который в течение нескольких опытов предшествовал мышечной работе. Усиление газообмена и увеличение легочной вентиляции, наблюдающееся еще до начала работы, способствуют большей выносливости и лучшей работоспособности организма во время мышечной деятельности. \ При сочетании условного раздражителя с дачей собаке пищи образуются наряду с двигательными условные секреторные рефлексы. Лучше и подробнее всего изучены слюноотделительные условные рефлексы. Все важнейшие закономерности условных реф- яексов получены лабораторией И. П. Павлова при экспериментальном анализе реакции слюноотделения. ' Секреторные компоненты условных пищевых рефлексов зарегистрированы также 1ри исследовании секреции желудка, и поджелудочной железы. Слюноотделительные условные рефлексы являются компонентами и оборонительного условного рефлекса, образующегося при сочетании условного сигнала с вливанием з рот собаке кислоты. Физиологическое значение этого, условного рефлекса состоит в удалении (смывании) раздражителя из полости рта. При исследовании условных рефлексов экспериментатор часто регистрирует какой- либо из его главных компонентов — соматический или вегетативный. В этом случае /словно говорят о двигательных, секреторных и вазомоторных рефлексах. Необходимо, эднако,учитывать, что они представляют собой лишь отдельные звенья целостной реакции организма. . Многочисленными и разнообразными экспериментами доказано, что в организме нет органа, деятельность которого не могла бы измениться в результате образования условного рефлекса. Любая функция целостного организма может быть усилена или заторможена под влиянием условнорефлекторных воздействий. Современные исследования показали широкие возможности регуляции функций, которыми располагает кора большого мозга. Условный рефлекс может образоваться и при сочетании условного сигнала с прямым электрическим или химическим раздражением коры полушарий большого мозга, или подкорковых ядер. Многократное, в течение 8—17 дней, впрыскивание морфина в одинаковых условиях и в одно и то же время вызывает образование условного рефлекса (появление слюноотделения, рвоты, шаткой походки, изменение дыхания) при действии одной обстановки опыта, при подготовке к впрыскиванию или при введении под кожу изотонического раствора хлорида натрия. После 10—12 сочетаний индифферентного раздражителя с подкожным введением бульбокапнина индифферентный раздражитель приобретал способность условнорефлек- торно воспроизводить картину бульбокапнинового отравления. У животного развивалось состояние каталепсии, при котором (так же как и при действии бульбокапнина) конечности становились воскоподобными, застывали в любом положении, так что собаке можно было придавать самые причудливые позы (А. И. Долин). ПРАВИЛА ОБРАЗОВАНИЯ УСЛОВНЫХ РЕФЛЕКСОВ И. П. Павлов и сотр. показали, что условный рефлекс можно выработать только тогда, когда начало действия индифферентного, т.е. не относящегося к данному виду деятельности, сигнала предшествует началу безусловного раздражения. Необходимо также, чтобы раздражитель, который мы хотим сделать условным, сам не вызывал значительной безусловной реакции, т.е. физическая сила (биологическая значимость) условного раздражителя не должна превышать силу (значимость) безусловного. Обязательными условиями выработки условных рефлексов являются нормальное деятельное состояние полушарий большого мозга, отсутствие патологических процессов в организме и каких-либо посторонних раздражений. МЕТОДИКА ИЗУЧЕНИЯ УСЛОВНЫХ РЕФЛЕКСОВ Для изучения условных рефлексов И.П. Павлов разработал оригинальную методику исследования. Используемый объект (животное или человек) изолируют от экспериментатора и ненужных по ходу опыта посторонних воздействий, помещая его в специальную камеру. Находясь вне камеры, экспериментатор на расстоянии включает раздражители и регистрирует изменения поведения испытуемого (рис. 234). 16* 483 Для изучения механизма образования условных рефлексов существенное значение имеёт не только точная регистрация самой ответной реакции (слюноотделения, движения и т.п.), но Также исследование электрической активности, возникающей в различных мозговых структурах во, время действия условного и безусловного раздражителей. Для регистрации электрической активности применяют электроды, хронически вживляемые в различные области или слои коры большого мозга, а также в специфические и неспецифические ядра таламуса, ретикулярную формацию, гиппокамп и другие отделы головного мозга. В опытах с условными рефлексами широко используются микроэлектродные методы, позволяющие регистрировать электрическую активность отдельных нейронов, участвующих в осуществлении условнорефлекторной реакции. Для автоматического анализа электроэнцефалограмм, регистрируемых с различных областей коры, в опытах на животных непосредственно во время условнорефлекторных реакций используют электронно-вычислительные машины. 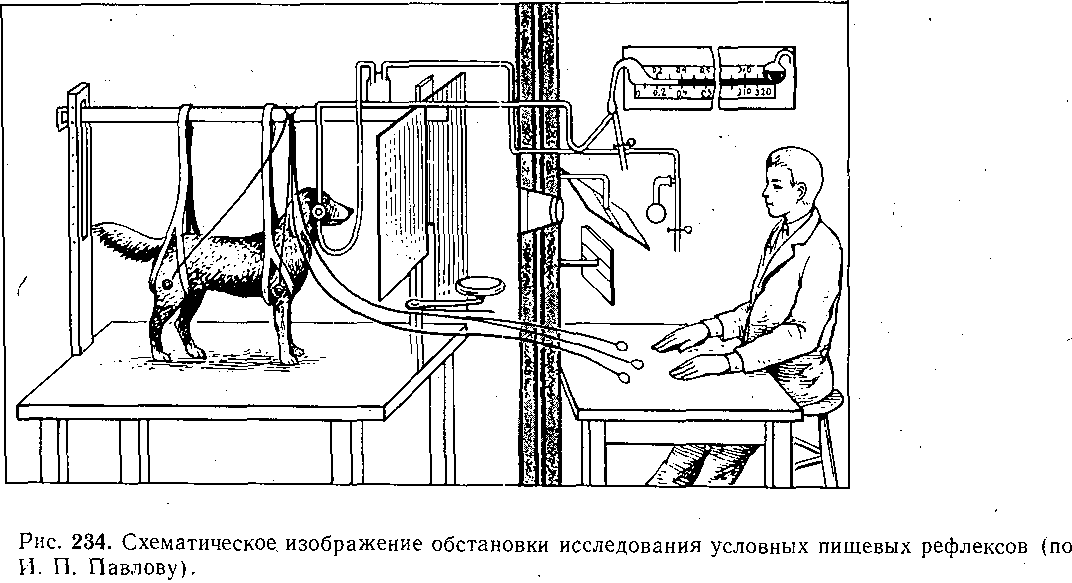 СИГНАЛЫ УСЛОВНОГО РЕФЛЕКСА Условным раздражителем может стать любое изменение окружающей среды или внутреннего состояния организма, достигшее определенной интенсивности и воспринятое корой полушарий большого мозга. Звуки (тоны и шумы), свет, контуры освещенных предметов, цвета, запахи, вкусовые средства, прикосновение к коже, давление, тепловые и холодовые воздействия, степень напряжения мышц (сокращение или расслабление)., положение тела в пространстве, состояние внутренних органов, воздействия на их слизистую оболочку, а также изменения обмена веществ и энергии в организме — все эти разнородные по своей природе раздражители при их сочетании с безусловными раздражениями становятся сигналами условных рефлексов. Следовые условные рефлексы образуются в том случае, если применяют безусловный раздражитель (например, пищевой) не во время действия условного сигнала, а лишь спустя определенное время после его окончания. В этом случае сам по себе сигнал не будет вызывать условного рефлекса, но после того, как он прекратится, возникает условнорефлекторное слюноотделение. Это означает, что именно след условного раздражителя в коре полушарий большого мозга приобрел для животного сигнальное значение. Если многократно каждые 10 мин кормить собаку, вырабатывается условный рефлекс, выражающийся в том, что к концу 10-й минуты после предыдущего кормления у животного начинает выделяться слюна и возникает двигательная реакция по направлению к кормушке. Условным сигналом, стал данный промежуток времени. Могут быть получены условные рефлексы и на гораздо более длительные периоды времени. Так, если ежедневно в определенный час кормить собаку, то к этому часу у нее еще до кормления начинается секреция желудочного сока. / При постоянном режиме труда и быта: точно определенных часах работы, приеме пищи в одно и то же время, одних и тех же часах сна — различные условные рефлексы на время наблюдаются и у человека. В естественной обстановке высшая нервная деятельность животных и поведение человека определяются не простыми световыми, звуковыми или иными одиночными сигналами, которыми пользуются в лаборатории, а всегда сложными, состоящими из комплекса одновременных звуковых, световых и других раздражителей. Например, для ребенка грудного возраста сигнал условного пищевого рефлекса — сосания — складывается из вида груди матери, звуков ее голоса, отчасти осязательных и обонятельных раздражений. Если сигналы разного рода следуют друг за другом всегда в определенном порядке, сопровождаясь в конце безусловным подкреплением, то образуется последовательный комплекс. В том случае, если при этом каждый следующий сигнал начинает свое действие только после окончания действия предыдущего, то формируется цепь раздражителей. Условные рефлексы на цепи раздражителей очень распространены. Например, обучение собаки выполнять приказ «дай лапу» складывается из цепи раздражителей: звукового («дай лапу»), проприоцептивного (поднятие конечности) и натурального пищевого (подкрепления пищей). ЗАВИСИМОСТЬ ВЕЛИЧИНЫ УСЛОВНОГО РЕФЛЕКСА ОТ СИЛЫ РАЗДРАЖИТЕЛЯ Величина условного рефлекса у животного при прочих равных условиях зависит как от силы безусловного рефлекса, на базе которого он вырабатывается, так от силы условного раздражителя. Если сочетать действие звукового агента с очень слабым электрокожным раздражением конечности собаки, то вырабатывающийся условный рефлекс оказывается слабым, неустойчивым. Если же силу безусловного раздражения увеличить, то это приведет к возникновению более сильного и стойкого оборонительного рефлекса. При постоянной силе безусловного раздражения величина условного, рефлекса зависит от физической силы сигнального раздражителя. Чем она больше, тем условный рефлекс сильнее (так называемый закон силовых соотношений). «Закон силы», однако, действителен только в определенных пределах, для всякого условного агента существует предел силы, за которым дальнейшее усиление раздражителя ведет к ослаблению условной реакции. УСЛОВНЫЕ РЕФЛЕКСЫ ВТОРОГО И ТРЕТЬЕГО ПОРЯДКА Условные рефлексы, образованные путем сочетания условного сигнала с безусловным раздражителем, получили название условных рефлексов первого порядка. Те условные рефлексы, которые образованы на основе сочетания внешнего агента с условным сигналом, вызывающим выработанный ранее прочный постоянный условный рефлекс первого порядка, называются условными рефлексами второго порядка.. Путем сочетания индифферентного раздражителя с условным раздражителем второго порядка можно выработать у собаки условный рефлекс третьего порядка. Последний удалось наблюдать в опытах с оборонительными двигательными рефлексами, которые вызывались электрическим раздражением лапы. Условных рефлексов четвертого порядка у собаки выработать не удается. У детей описаны рефлексы шестого порядка. Изучение природы временной связи условного рефлекса открывает путь к познанию основы высших функций мозга — явлений памяти. Однако несмотря на многочисленные исследования, до сих пор очень многое остается неясным. Структура условного рефлекса. Временная связь, лежащая в основе условного рефлекса, имеет «горизонтальную» структуру и замыкается в пределах коры между центром сигнального раздражителя и клетками коркового представительства безусловного рефлекса. Временные связи образуются, как правило, на корковом уровне, однако при исключении такой возможности, например при разрезах коры, могут осуществляться по путям нижележащих уровней (рис. 235, I). Многие особенности выработки условных рефлексов получают объяснение, если принять, что при этом формируются множественные прямые и обратные связи как в пределах коры, так и между корой и подкорковыми центрами (рис. 235, II). Разрез 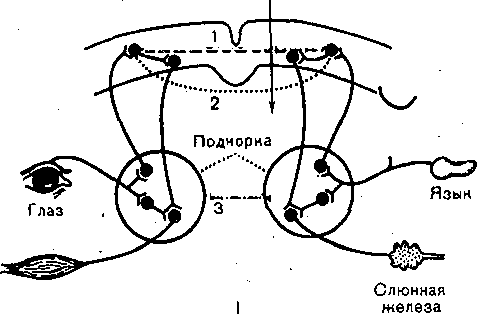 Мышца 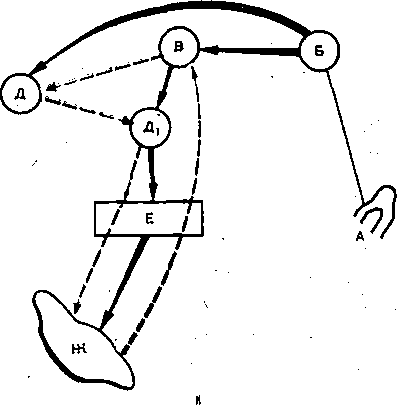 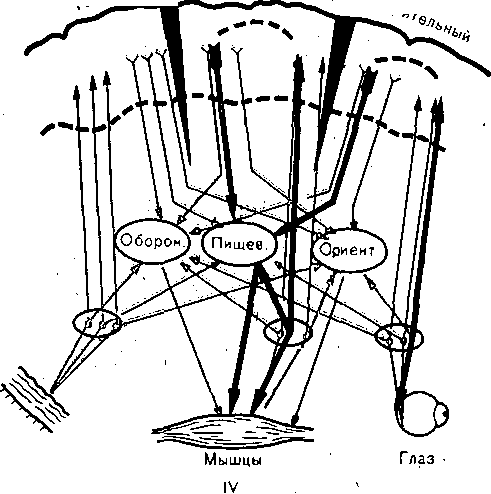 а. За Ножа 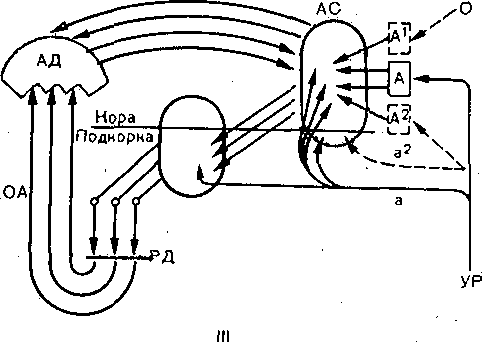 Рис. 235. Гипотезы формирования условного рефлекса (схемы). 1 — временная связь между зрительным и слюноотделительным безусловным рефлексом (по Э. А. Асратяну).
А — слуховые рецепторы; Б, В — слуховая проекционная область; Д — пищевой центр; Е — двигательные элементы коры; Ж — подкорковые двигательные аппараты.
УР — условный раздражитель; а — неспецифические и а2 — специфические афференты; О — раздражители обстановки; А, А1, Аг — анализаторы обстановочных и пусковых сигналов; АС — аппарат афферентного синтеза; АД — аппарат акцепторного действия; РД — рефлекторное действие; OA — обратная афферента- ция.
Изучение функциональной организации условного рефлекса привело к представлению о формировании в коре и подкорковых центрах аппарата афферентного синтеза, акцептора результатов действия и обратной афферентации о результатах (рис. 235, III). Возможно, что структурная пластичность временных связей, которым не препятствует рассечение коры в любом месте, обеспечивается ее способностью замыкать эти связи в каждом своем участке между афферентными и эфферентными клетками, осуществляя «распределенную память» (рис. 235, IV). Определение локализации замыкания временной связи условного рефлекса по параметрам реакций фоновой электрической активности различных структур мозга затрудняется тем, что данные реакции, с одной стороны, имекл чрезвычайно обширный характер за счет своих неспецифических компонентов, а с дру гой — по мере укрепления условного рефлекса, как правило, идут на убыль, чем такж* подтверждается их неспецифичность. Вместе с тем описаны изменения некоторых позд них компонентов вызванных потенциалов, возникающих в ответ на условные и безуслов ные раздражители в зонах корковых проекций, которые оказываются более стабильными и могли бы указывать на более близкое отношение этих зон коры к структуре временно* связи условного рефлекса. Высказывалось мнение, что замыкание временной связи осуществляют главныл образом вставочные и ассоциативные нейроны коры большого мозга (И. С. Беритов) Каждый вид корковой деятельности складывается из взаимодействия множества возбуж дающихся и тормозящихся нейронов. Поэтому изучение структуры временной связи н< нейронном уровне представляет сложную задачу, которая еще далеко не решена. МЕХАНИЗМЫ ЗАМЫКАНИЯ ВРЕМЕННОЙ СВЯЗИ До настоящего времени нет единой точки зрения на механизмы замыкания временных связей между нейронами высших отделов ЦНС в процессе образования условны* рефлексов. Морфологическое объяснение, выдвинутое еще в прошлом веке, предполагало, что во время образования условного рефлекса происходит рост отростков нервны* клеток, устанавливающих новые межклеточные связи. Сходные идеи высказывались v позже, в частности относительно роли своеобразных выростов — «шипиков» на апикаль ных дендритах пирамидных нейронов. Функциональное объяснение видело ответ на поставленный вопрос в повышенной проводимости существующих синапсов, которая может обусловливаться непрерывной циркуляцией по нейронным кругам, увеличением числа используемых синапсов, т.е. явлениями, подобными посттетанической потенциации. Было высказано предположение и о возможной роли глии в организации условно- рефлекторной деятельности. В настоящее время накопилось много фактов, свидетельствующих о том, что функция глиальной ткани не сводится лишь к опорной и трофической Возможность участия глиальных клеток в образовании стойкой временной связи заключается в миелинизации «голых» пресинаптических терминалей аксонов, что делает ю способными быстро проводить импульсы. Многие исследователи считают ведущим фактором изменения синтеза белка в клетках. Исследование нуклеотидного состава рибонуклеиновой кислоты (РНК) в процесс* выработки условных рефлексов позволило предположить, что совместное возбуждение многих нейронов отражается на структуре РНК, в которой кодируется сигнальное разд ражение. В свете этой гипотезы трактовались результаты экспериментов, показавши? облегчение выработки новых навыков у червей, которым скормили тех, у кого этот навык был заранее выработан. Такой «перенос химической памяти» был описан рядом исследователей на основе опытов с введением различным животным экстрактов из мозга животных, обученных какому-либо навыку и его устранение при действии фермента рибонуклеазы, разрушающей РНК (ряд исследователей не подтвердили эти результаты). Некоторые считают более вероятным, что след памяти закрепляется на молекулах дезоксирибонуклеиновой кислоты (ДНК), которая сохраняет этот след дольше, чем относительно быстро синтезируемая РНК. Изменение свойств ДНК и РНК реализуется через количественные и качественные изменения синтеза белков. Описаны специфические белковые вещества мозга, с которыми связывают фиксацию временной связи между нейронами вплоть до попыток выделения пол и пептидных комплексов, ответственных за воспроизведение оборонительных, пищевых и других условных рефлексов. В других гипотезах на первый план в механизмах формирования временной связи выдвигаются системы белков-ферментов и их ингибиторов, которые могут изменять свое состояние. i i I i i t i , i i '' i j ФВ 100 200 300 мим ФВ 100 200 300 мкм ' а б >ис. 236. Распределение возбуждающихся (В), тормозящихся (Т) и нереагирующих (Н) нейронов рительной коры мозга морской свинки в фокусе возбуждения (ФВ) и на разном удалении от него по А. Б. Когану и Подладчиковой). а — первичные реакции 502 нейронов; б -— вторичные реакции 493 нейронов. На ординате — процентные доли нейронов В, Т, Н; на абсциссе — расстояние от фокуса возбуждения. Переход от-изучения деятельности отдельных нейронов к изучению их взаимодей- :твия приводит к трактовке механизма временной связи условного рефлекса как форми- ювания определенного узора нейронной активности на основе избирательной общности :войств реагирования. Возникновение узора определяется такими условиями, которые )блегчают проведение возбуждения в структурах мозга: синхронность и синфазность 1ктивн0сти в определенном диапазоне частот, (переход значительного числа нейронов, >бразующих цепочки возбуждения, на сходные уровни лабильности). Взаимодействие >чагов возбуждения, ограниченных торможением, которое формируется под влиянием 1фферентных сигналов, имеет динамический характер и наиболее выражено в фазу пер- $ичной активации нейронов (рис. 236). За последнее время представление о временной :вязи как пространственно-временной организации межнейронных отношений (получив- цей название нейроселективного действия) подкрепляется и нейрохимическими иссле- ^ваниями, которые позволили сделать заключение о специфическом изменении химизма lefipoHOB, объединяющем их в функциональные ансамбли, об избирательной активации 1ерез ДНК нейроглиальных комплексов. ч. ПАМЯТЬ Временная нервная связь, лежащая в основе образования условного рефлекса, — шшь частный случай общебиологического свойства хранения воспринятой информации, шторое в той или иной степени проявляется у всех живых существ, начиная с простей- пих. Временную связь оказалось возможным выработать у растений, например, на базе защитной )еакции стыдливой мимозы или суточного цикла складывания листьев у .бобовых. В широком смысле такое свойство, обеспечивающее запечатлевание связей событий жружающего мира, что позволяет накапливать и использовать жизненный опыт, является памятью. О механизме возникающих при этом межклеточных временных связей пока лало известно. Рис. 237. Определение времени консолидации следа долгосрочной памяти при образовании оборонительного условного рефлекса (задержки выхода мыши на платформу), за которым следовал удар тока. На ординате — задержка выхода на платформу в секундах, на абсциссе — время в минутах от сочетания выхода с ударом тока до воздействия кратковременным эфирным наркозом. Белые столбики — выработка рефлекса без наркотизации, заштрихованные — после наркотизации в разные сроки. В нервной системе механизмы памяти получили чрезвычайное развитие и приобрели ведущую роль в поведении. По проявлениям различают память образную, которая воспроизводит образ жизненно важного объекта, эмоциональную, когда аналогичная ситуация вызывает эмоции, сопровождавшие происходившие ранее в этой ситуации события, и словесно-логическую, свойственную только человеку (И. С. Беритов). По времени сохранения различают кратковременную и долговременную память, имеющие в своей основе разные механизмы. Кратковременную память называют также оперативной, так как она обеспечивает выполнение текущих операций мышления (например, умножая 26 на 4, пишем 4, а 2 «держим в уме», чтобы через несколько секунд прибавить к 8). Предполагают, что механизмом кратковременной памяти может быть циркуляция импульсных потоков по замкнутым кругам нейронных цепей. Этому соответствует то обстоятельство, что кратковременная память легко нарушается под действием сильных посторонних раздражений, наркозом, электрической стимуляцией мозга, его гипоксией, различными нейротропными ядами, которые при той же интенсивности действия не затрагивают память долговременную. В основе же долговременной памяти лежат, по-видимому, сложные процессы, связанные с активностью синтеза белковых молекул в клетках большого мозга. ' Как показано на рис. 237, время, необходимое для перехода кратковременной памяти в долговременную, в условиях эксперимента составляет около 30 с. Однако существует мнение, что речь идет лишь о последовательных этапах консолидации единого механизма памяти, а не о двух ее разновидностях. Сопоставление обширного материала клинических наблюдений и результатов разнообразных экспериментов на животных позволяет считать, что память как единый процесс складывается по крайней мере из трех взаимосвязанных этапов, возможно, имеющих'различные механизмы: запоминания, хранения опыта и воспоминания (воспроизведение опыта). ТОРМОЖЕНИЕ УСЛОВНЫХ РЕФЛЕКСОВ БЕЗУСЛОВНОЕ ТОРМОЖЕНИЕ При действии разнообразных посторонних раздражителей условные рефлексы легко подвергаются торможению. Так, если перед началом или во время условного пищевого рефлекса внезапно возникает посторонний звук, появляется какой-либо посторонний запах, резко изменяется освещение и т.п., то условный рефлекс ослабляется или исчезает. Объясняется это тем, что всякий новый раздражитель вызывает у собаки ориентировочный рефлекс, который тормозит условную реакцию. При многократном повторении такого раздражителя ориентировочная реакция на него угасает и ослабляется его тормозящее действие на условный рефлекс. Подобные посторонние раздражители И. П. Павлов назвал «гаснущими тормозами». 30,0 25,0 20,0 - 15,0 10,0 5,0 ш I I I 6 8 10 12 14 16 18 20 22 24 Г71 Р71 Г71 > ' ' ' г ■ I Более устойчивый тормозящий эффект на условные рефлексы дают раздражители, вызывающие посторонние (по отношению к данному) безусловные или условные рефлек торные реакции. Например, болевое раздражение или сигнал о нем резко тормозит пищевые условные рефлексы. Так же могут действовать и раздражения, исходящие от внутренних органов. Переполнение мочевого пузыря, рвота, половое возбуждение, воспалительный процесс в каком-либо органе вызывают угнетение условных пищевых рефлексов. Все эти случаи торможения имеют между собой общее: они возникают под влиянием внешних, посторонних для данного условного рефлекса раздражений. Поэтому такое торможение называли внешним торможением. |